宿主細(xì)胞的脂筏在問號鉤端螺旋體的進(jìn)入和存活中起關(guān)鍵作用
2021-06-17 12:56:46呂天寶王佳琪謝旭峰關(guān)鈺穎吳殿君張文龍曹永國
中國獸醫(yī)學(xué)報 2021年5期
呂天寶,王佳琪,謝旭峰,宋 寧,關(guān)鈺穎,吳殿君,張文龍,曹永國
(吉林大學(xué) 動物醫(yī)學(xué)學(xué)院, 吉林 長春 130062)
鉤端螺旋體病是一種由致病性鉤端螺旋體(鉤體)引起的,除南極洲以外世界范圍內(nèi)廣泛存在的人獸共患病[1]。鉤體是一種專性需氧的革蘭陰性細(xì)菌,可分為非致病性和致病性兩大類,呈螺旋形,長6~20 μm,直徑約0.1 μm[1]。鉤體通過擦傷、割傷或直接穿過正常的皮膚和黏膜進(jìn)入人體,然后進(jìn)入血管并通過血流到達(dá)全身組織器官,最終定居在腎臟,鉤體經(jīng)患病動物或人的尿液排出,污染水源和土壤,人和動物可因直接或間接接觸被污染的水和土壤而感染[2]。不同的動物感染不同血清型鉤體表現(xiàn)出不同的臨床癥狀,從輕度的流感樣疾病到嚴(yán)重的感染,包括腎和肝衰竭,肺部疾病和死亡。典型癥狀為黃疸,典型病理變化為肺出血[1]。迄今為止,有關(guān)鉤體入胞機(jī)制尚未被闡明。
脂筏是質(zhì)膜中功能性的納米級膜結(jié)構(gòu)域,富含膽固醇和鞘脂,其中包含多種信號傳導(dǎo)和轉(zhuǎn)運蛋白[3]。越來越多的病原體(包括細(xì)菌、病毒和寄生蟲等)如大腸埃希菌、假單胞菌、衣原體和A型流感病毒等,已被證明其侵入宿主細(xì)胞需要脂筏[4-7]。例如,SEVEAU等[8]發(fā)現(xiàn)鈣黏蛋白和InlB-HGF-R/Met介導(dǎo)的單核細(xì)胞增生性李斯特菌進(jìn)入細(xì)胞需要脂筏結(jié)構(gòu)的完整性。但是,鉤體進(jìn)入細(xì)胞是否與脂筏相關(guān)尚未見報道。
1 材料與方法
1.1 細(xì)菌和細(xì)胞致病性問號鉤端螺旋體賴型賴株(56601)、鼠巨噬細(xì)胞細(xì)胞系(J774A.1,ATCC編號TIB-67)和非洲綠猴腎上皮細(xì)胞(Vero細(xì)胞,ATCC編號CCL-81)由本實驗室保存。
1.2 主要儀器與試劑CO2細(xì)胞培養(yǎng)恒溫箱購自日本三洋公司;暗視野顯微鏡購自德國奧林巴斯公司;Petroff-Hauser細(xì)菌計數(shù)板購自美國托馬斯公司;共聚焦顯微鏡購自德國奧林巴斯公司;通用型電泳儀購自美國Bio-Red公司;DNA提取試劑盒購自天根生化科技(北京)有限公司;熒光定量試劑盒購自美國羅氏公司;LC3B、LAMP-1和Caveolin-1抗體購自美國Abcam公司;β-actin以及各種二抗購自美國Immunoway公司;MβCD和膽固醇購自美國Sigma公司;3MA和雷帕霉素購自中國瀚香生物科技有限公司;Lipofectamine RNAiMAX試劑購自美國Invitrogen公司;Cav-1 siRNA購自Santa Cruz Biotechnology公司。
1.3 細(xì)胞與細(xì)菌培養(yǎng)致病性問號鉤端螺旋體賴型賴株(56601)在29℃的Ellinghausen-McCullough-Johnson-Harris(EMJH)液體培養(yǎng)基中培養(yǎng),通過感染金黃地鼠保持毒力。對于所有細(xì)胞試驗,鉤體在液體EMJH培養(yǎng)基中的傳代次數(shù)少于3次。J774A.1和Vero細(xì)胞分別培養(yǎng)在含有10%和2%胎牛血清、100 U/mL青霉素和100 mg/L鏈霉素的RPMI 1640培養(yǎng)基中,在37℃、5% CO2細(xì)胞培養(yǎng)箱中培養(yǎng)。
1.4 細(xì)胞毒性試驗將J774A.1細(xì)胞以5×103/孔的密度接種在96孔板中,待細(xì)胞貼壁后加入不同濃度的MβCD作用30 min后,用CCK8試劑盒檢測細(xì)胞毒性。
1.5 細(xì)胞處理與感染將J774A.1細(xì)胞和Vero細(xì)胞以1×106個/孔的密度接種在6孔板中,在完全培養(yǎng)基中預(yù)孵育12 h。將細(xì)胞分為3組:對照組、MβCD組和MβCD+膽固醇組,然后將MβCD組在含有(10 mmol/L)MβCD的RPMI 1640培養(yǎng)基中于37℃孵育30 min,MβCD+膽固醇組在含有10 mmol/L MβCD的RPMI 1640培養(yǎng)基中處理30 min 后再在含200 mg/L膽固醇的RPMI 1640培養(yǎng)基中37℃孵育30 min。用PBS洗滌后,3組細(xì)胞以1∶100的劑量感染致病性菌株56601。將細(xì)胞在5% CO2中于37℃孵育40 min,用PBS洗滌,經(jīng)胰蛋白酶消化的離心(1 200 r/min)收集細(xì)胞,使用DNA提取試劑盒提取DNA以測量胞內(nèi)鉤體量。
將J774A.1細(xì)胞以1×106個/孔的密度接種在6孔板中,12 h后以1∶100的比例感染鉤體,同時加入3MA(10 mmol/L)或雷帕霉素(50 mg/L),2 h后收集細(xì)胞,用1 mL蒸餾水裂解細(xì)胞,吸取100 μL裂解液加入到3 mL EMJH培養(yǎng)基中,29℃培養(yǎng)箱中恒溫培養(yǎng)4~6 d,在暗視野顯微鏡下用Petroff-Hauser細(xì)菌計數(shù)板計數(shù)。
1.6 小窩蛋白(Cav-1)的小RNA干擾試驗使用Lipofectamine RNAiMAX試劑將siRNA轉(zhuǎn)染到J774A.1細(xì)胞中,并在24 h后對Cav-1進(jìn)行免疫熒光染色,對細(xì)胞進(jìn)行鉤體感染試驗。
1.7 實時熒光定量PCR根據(jù)致病性鉤體特異性基因LipL32設(shè)計引物[9],對胞內(nèi)鉤體進(jìn)行熒光定量PCR分析(絕對定量),使用體外培養(yǎng)細(xì)菌中提取的DNA的梯度稀釋液(109~102)制備標(biāo)準(zhǔn)曲線,測定鉤體量。
1.8 激光共聚焦顯微鏡將多聚賴氨酸處理過的12 mm細(xì)胞爬片置于24孔板中,以2×105個/孔的數(shù)量接種細(xì)胞,待細(xì)胞貼壁后,將細(xì)胞分為對照組和MβCD組,分別處理后,以1∶100的比例感染鉤體,感染40 min后,吸去上清,用預(yù)冷的PBS清理3次,加4%的多聚甲醛固定15 min,PBS清洗3次后,以含2% BSA的PBS溶液室溫封閉30 min,用抗鉤體一抗室溫孵育30 min,PBS清洗3次后,用FITC標(biāo)記的熒光二抗室溫孵育30 min,對細(xì)胞外的鉤體進(jìn)行熒光染色。PBS清洗3次后,用含0.2% Triton X-100的PBS溶液室溫處理10 min,PBS清洗1次后再用含2% BSA的PBS溶液室溫封閉30 min,用抗鉤體一抗室溫孵育30 min,PBS清洗3次后,用TRITC熒光二抗室溫孵育30 min,對胞內(nèi)鉤體進(jìn)行熒光染色。用PBS清洗3次后,用Hoechst染色5 min,清洗3次后將細(xì)胞爬片以甘油黏附到載玻片上,置于共聚焦顯微鏡下觀察。
1.9 Western blot將J774A.1細(xì)胞以1×106個/孔的密度接種在6孔板中,并在完全培養(yǎng)基中預(yù)孵育12 h。將細(xì)胞分為4組:對照組、MβCD組、鉤體組和MβCD+鉤體組。用MβCD刺激30 min后感染細(xì)胞,于感染后4 h收集細(xì)胞,用預(yù)冷的PBS緩沖液清洗3遍,用蛋白提取試劑裂解細(xì)胞10 min,然后用超速離心機(jī)以4℃ 12 000 r/min離心10 min收集上清液,使用BCA法檢測蛋白濃度。調(diào)整蛋白濃度后加入蛋白上樣緩沖液,100℃加熱8 min使蛋白變性,配置15% SDS-PAGE濃縮膠以及分離膠,以30 μg/孔蛋白量上樣,80 V電泳20 min后轉(zhuǎn)用120 V 電泳1 h,然后通過濕轉(zhuǎn)法將蛋白轉(zhuǎn)至PVDF膜上,用5%的脫脂奶粉封閉2 h,用相應(yīng)的一抗在4℃環(huán)境下孵育過夜,用TBST溶液清洗3次后,常溫下用二抗孵育2 h,用TBST溶液清洗3次后即可在顯影儀下檢測蛋白量。
2 結(jié)果
2.1 鉤體進(jìn)入細(xì)胞需要脂筏介導(dǎo)免疫熒光和Western blot結(jié)果表明,MβCD和Cav-1 siRNA都能破壞細(xì)胞的脂筏結(jié)構(gòu)(圖1),并且,細(xì)胞毒性試驗分析表明MβCD對細(xì)胞沒有潛在的毒性(圖2)。因此,MβCD和Cav-1 siRNA能夠用做脂筏破壞的試劑進(jìn)行接下來的試驗。
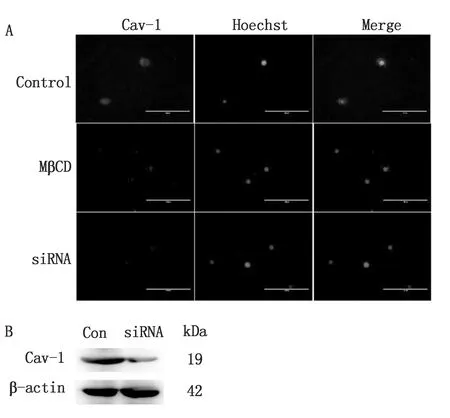
A.MβCD(10 mmol/L)刺激細(xì)胞30 min以及Cav-1 siRNA(100 nmol/L)刺激細(xì)胞24 h后,對細(xì)胞小窩蛋白的熒光染色,其中每個樣品至少隨機(jī)拍攝10張不同部位的圖片,圖中展示了各組代表性結(jié)果(×400);B.Western blot檢測Cav-1 siRNA的沉默效果,試驗重復(fù)3次
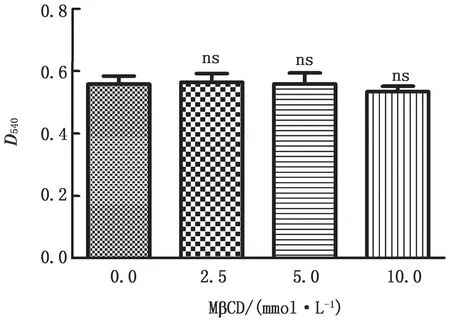
以不同濃度MβCD處理J774A.1細(xì)胞30 min,然后用CCK8試劑盒檢測細(xì)胞毒性。ns表示P>0.05,無顯著性差異
用qPCR檢測細(xì)胞內(nèi)的鉤體量,結(jié)果表明,用MβCD破壞脂筏后,進(jìn)入J774A.1細(xì)胞內(nèi)的鉤體量明顯降低,用膽固醇回補脂筏后,胞內(nèi)鉤體量有所升高(圖3A),證明鉤體進(jìn)入細(xì)胞需要脂筏。為了驗證這個現(xiàn)象,采用了小RNA干擾技術(shù),敲低了細(xì)胞的小窩蛋白,結(jié)果表明,敲低細(xì)胞的小窩蛋白后,進(jìn)入J774A.1細(xì)胞的鉤體量明顯降低(圖3C)。在Vero細(xì)胞的試驗中,也發(fā)現(xiàn)了同樣的現(xiàn)象,即用MβCD破壞脂筏后,細(xì)胞內(nèi)的鉤體量明顯降低,用膽固醇回補脂筏后,胞內(nèi)鉤體量有所升高(圖3B)。激光共聚焦結(jié)果直觀地表明,脂筏被破壞后,胞內(nèi)的鉤體量明顯減少(圖3D)。
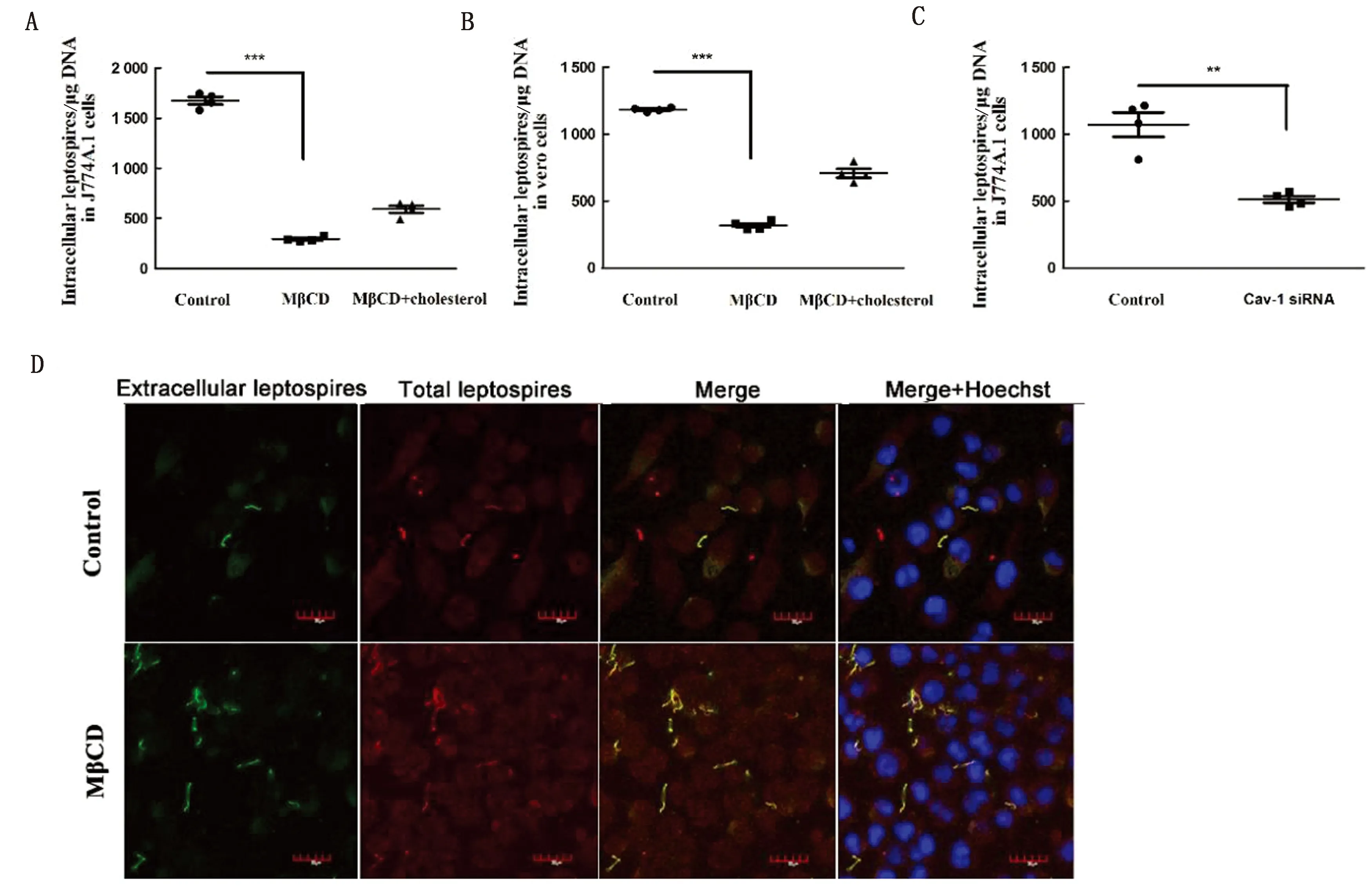
A,B.J774A.1和Vero細(xì)胞在正常狀態(tài),MβCD破壞脂筏的狀態(tài)以及膽固醇回補狀態(tài)下,鉤體進(jìn)入細(xì)胞的數(shù)量的檢測,試驗重復(fù)3次;C.J774A.1細(xì)胞在正常狀態(tài)以及Cav-1 siRNA破壞脂筏的狀態(tài)下,進(jìn)入細(xì)胞的鉤體數(shù)量檢測,試驗重復(fù)3次;D.激光共聚焦顯微鏡下觀察胞內(nèi)外鉤體的熒光染色結(jié)果(×1 000)。*表示P<0.05,顯示有顯著性差異;**表示P<0.01,顯示有非常極顯著性差異;***表示P<0.001,顯示有極顯著性差異。下同
2.2 脂筏影響胞內(nèi)鉤體的存活透射電子顯微鏡下觀察在有無脂筏的情況下鉤體進(jìn)入細(xì)胞的情況,結(jié)果顯示在感染后40 min,破壞脂筏結(jié)構(gòu)的J774A.1細(xì)胞(圖4E)比正常細(xì)胞(圖4B)中的鉤體量明顯較少。在感染后4 h,正常細(xì)胞中還有大量鉤體,其形態(tài)學(xué)結(jié)構(gòu)無明顯改變(圖4C),但是在脂筏被破壞的細(xì)胞中,感染后4 h的胞內(nèi)鉤體基本被殺死,并且形態(tài)學(xué)結(jié)構(gòu)模糊(圖4F)。另外,通過qPCR檢測了感染后4 h的胞內(nèi)鉤體量,結(jié)果表明,與正常組相比,脂筏被破壞后,細(xì)胞殺滅鉤體的能力增強(qiáng)(圖4G)。此外,液體培養(yǎng)基分離培養(yǎng)試驗也證明了脂筏被破壞后,細(xì)胞殺滅鉤體的能力增強(qiáng)(圖4H)。
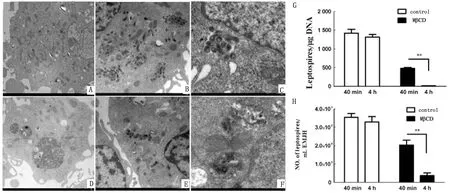
A.透射電子顯微鏡下,正常狀態(tài)下J774A.1細(xì)胞圖;B.正常J774A.1細(xì)胞感染后40 min胞內(nèi)鉤體的情況;C.正常J774A.1細(xì)胞感染后4 h胞內(nèi)鉤體的情況;D.脂筏破壞后的J774A.1細(xì)胞圖;E.脂筏破壞后,J774A.1細(xì)胞感染后40 min胞內(nèi)鉤體的情況;F.脂筏破壞后,J774A.1細(xì)胞感染后4 h胞內(nèi)鉤體的情況;G.qPCR檢測感染后40 min以及4 h J774A.1細(xì)胞在有無脂筏狀態(tài)下胞內(nèi)的鉤體量,試驗重復(fù)3次;H.液體培養(yǎng)基培養(yǎng)計數(shù)結(jié)果表示感染后40 min以及4 h J774A.1細(xì)胞在有無脂筏狀態(tài)下胞內(nèi)存活的鉤體量,試驗重復(fù)3次。
2.3 破壞脂筏增強(qiáng)了鉤體感染引起的細(xì)胞自噬水平Western blot結(jié)果表明,用MβCD破壞脂筏后,J774A.1細(xì)胞LC3-Ⅱ蛋白的水平明顯增高,溶酶體標(biāo)記性蛋白LAMP-1的水平也明顯增高(圖5A)。并且,在感染鉤體后,與對照組相比,脂筏破壞組的LC3-Ⅱ和LAMP-1蛋白的水平明顯較高。證明脂筏被破壞,感染鉤體后,細(xì)胞的自噬水平明顯增強(qiáng)(圖5A)。另一方面,用自噬的抑制劑(3MA)和激活劑(雷帕霉素)分別刺激感染鉤體的J774A.1細(xì)胞,用液體培養(yǎng)基培養(yǎng)計數(shù)方法測定胞內(nèi)鉤體量,結(jié)果表明,用雷帕霉素激活細(xì)胞自噬后,胞內(nèi)存活的鉤體明顯較少(圖5B)。由以上結(jié)果可知,破壞脂筏可以增強(qiáng)細(xì)胞自噬水平,自噬水平的增強(qiáng)有助于細(xì)胞殺滅胞內(nèi)的鉤體。
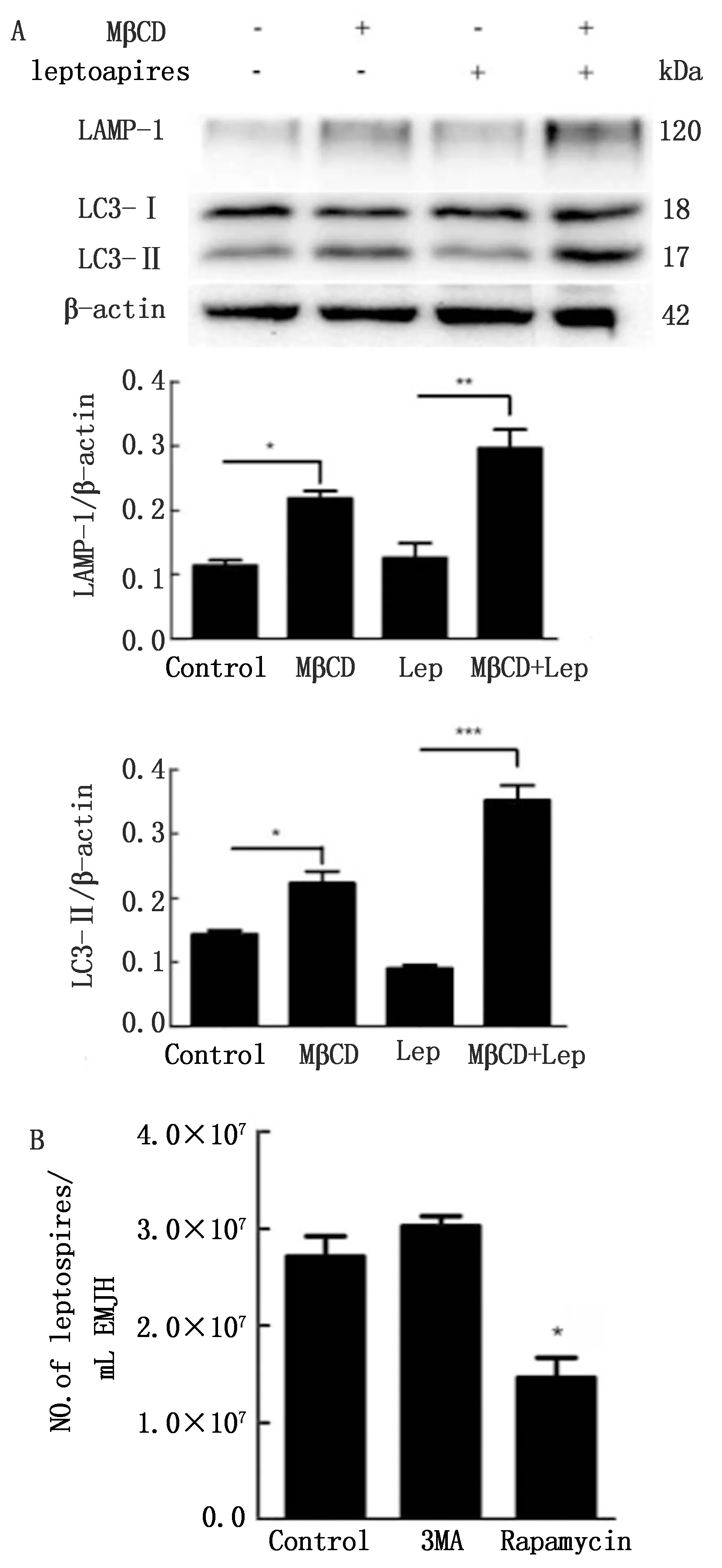
A.脂筏完整和被破壞狀態(tài)下感染鉤體后4 h的自噬標(biāo)記分子蛋白免疫印跡圖以及LC3-Ⅱ和LAMP-1蛋白的相對表達(dá)水平,試驗重復(fù)3次;B.3MA和雷帕霉素分別刺激細(xì)胞后,用液體培養(yǎng)基培養(yǎng)計數(shù)測得細(xì)胞內(nèi)存活的鉤體量
3 討論
鉤體為非專性細(xì)胞內(nèi)寄生菌,能夠在哺乳動物細(xì)胞內(nèi)轉(zhuǎn)運而不損壞細(xì)胞的功能。鉤體通過人或動物破損的皮膚或黏膜進(jìn)入體內(nèi),隨血流遍布全身組織器官。敏感動物感染后急性死亡,耐受動物發(fā)展成慢性腎臟疾病,其腎臟中有大量鉤體定殖,并隨尿液排出,污染土壤和水源,嚴(yán)重威脅著社會公共衛(wèi)生安全[10]。我國農(nóng)業(yè)部第1125號公告也將鉤體病列為我國二類動物疫病,第1149號公告將其列入人獸共患傳染病名錄。但是,鉤體進(jìn)入細(xì)胞的機(jī)制以及其在胞內(nèi)移行及胞間的穿行機(jī)制還未被闡明。
脂筏是脂膜上富含鞘磷脂和膽固醇的微結(jié)構(gòu)域,其包含多種信號轉(zhuǎn)導(dǎo)蛋白和轉(zhuǎn)運蛋白,例如小窩蛋白[11-12]。越來越多的病原體,例如大腸埃希菌、假單胞菌、衣原體和A型流感病毒等,已被證明其侵入宿主細(xì)胞需要脂筏[4-7]。因此,需要探究脂筏是否在鉤體入胞過程中發(fā)揮作用。
試驗證明MβCD對細(xì)胞無毒性,并且免疫熒光結(jié)果表明,J774A.1細(xì)胞在MβCD作用30 min后,脂筏的標(biāo)志性蛋白小窩蛋白1(Caveolin-1)的表達(dá)明顯降低,因此,參照之前的研究[13-14],采用MβCD破壞細(xì)胞的脂筏結(jié)構(gòu)。與對照組相比,在對J774A.1細(xì)胞進(jìn)行脂筏結(jié)構(gòu)的破壞后,進(jìn)入細(xì)胞的鉤體量明顯降低,用膽固醇回補脂筏結(jié)構(gòu),發(fā)現(xiàn)胞內(nèi)的鉤體量有所升高,證明鉤體進(jìn)入J774A.1細(xì)胞需要脂筏。為了避免單一藥物的偶然性,用小RNA干擾技術(shù)敲低細(xì)胞Cav-1的表達(dá)[15],結(jié)果表明,Cav-1的表達(dá)降低后,進(jìn)入細(xì)胞的鉤體量同樣減少,再次驗證了鉤體進(jìn)入細(xì)胞需要脂筏。為了驗證這個現(xiàn)象是否與細(xì)胞類型有關(guān),用Vero細(xì)胞重復(fù)了同樣的試驗,結(jié)果也表明鉤體進(jìn)入細(xì)胞對脂筏的依賴性。另外,激光共聚焦顯微鏡的觀察結(jié)果更直觀地顯示這一現(xiàn)象。這些結(jié)果共同表明鉤體進(jìn)入細(xì)胞需要脂筏,當(dāng)脂筏結(jié)構(gòu)的完整性被破壞后,鉤體進(jìn)入細(xì)胞的能力明顯降低,并且,這個現(xiàn)象與巨噬細(xì)胞的主動吞噬作用無關(guān)。這種對脂筏的依賴性在單核細(xì)胞增生性李斯特桿菌和豬流行性腹瀉病毒的入胞過程中同樣被證實[8,16]。
接下來,通過透射電子顯微鏡觀察J774A.1細(xì)胞在有無脂筏結(jié)構(gòu)的情況下感染后40 min及感染4 h后胞內(nèi)鉤體的數(shù)量和形態(tài)。結(jié)果表明,在感染后40 min,破壞脂筏結(jié)構(gòu)的J774A.1細(xì)胞內(nèi)的鉤體量明顯減少;在感染后4 h,對照組胞內(nèi)仍有大量鉤體,并且其形態(tài)完整,而脂筏破壞組胞內(nèi)的鉤體基本消失,形態(tài)模糊。從胞內(nèi)鉤體全菌熒光定量和活菌的液態(tài)培養(yǎng)基培養(yǎng)計數(shù)結(jié)果可知,在感染后4 h,脂筏破壞組胞內(nèi)的鉤體幾乎全被清除。這些結(jié)果表明破壞脂筏后,細(xì)胞殺滅鉤體的能力增強(qiáng)。
有研究表明,脂筏AKT-FOXO1途徑會影響自噬的調(diào)節(jié),脂筏的破壞會誘導(dǎo)自噬[17]。細(xì)胞自噬是真核細(xì)胞中通過溶酶體降解細(xì)胞質(zhì)成分的過程,同時也具有重要的宿主防御功能。因此,探究中破壞脂筏后,細(xì)胞殺滅鉤體的能力增強(qiáng)是否與細(xì)胞自噬增強(qiáng)有關(guān),結(jié)果表明,破壞脂筏能夠顯著增強(qiáng)細(xì)胞LC3-Ⅱ和LAMP-1蛋白表達(dá),證明破壞脂筏能夠增強(qiáng)自噬水平以及鉤體感染后的自噬水平。因此,細(xì)胞自噬水平的升高可能有助于殺滅胞內(nèi)的鉤體。為驗證這一猜想,分別用自噬激活劑(雷帕霉素)和自噬抑制劑(3MA)處理鉤體感染的J774A.1細(xì)胞,液體培養(yǎng)基培養(yǎng)計數(shù)結(jié)果表明,雷帕霉素能夠顯著降低胞內(nèi)鉤體量,證明自噬水平的增強(qiáng)有助于殺滅胞內(nèi)鉤體。以上結(jié)果表明,破壞細(xì)胞脂筏增強(qiáng)了細(xì)胞自噬和細(xì)胞殺滅鉤體的能力。
綜上所述,本研究證明鉤體進(jìn)入細(xì)胞需要脂筏,在破壞脂筏后,鉤體進(jìn)入細(xì)胞的量明顯降低,并且細(xì)胞殺滅鉤體的能力增強(qiáng),可能的原因是因為破壞脂筏加強(qiáng)了細(xì)胞的自噬水平。本研究發(fā)現(xiàn)鉤體進(jìn)入細(xì)胞與脂筏結(jié)構(gòu)的完整性相關(guān),為鉤體病預(yù)防產(chǎn)品的研發(fā)提供理論依據(jù)。
