布魯菌Ⅳ型分泌系統非依賴型效應蛋白BspI誘導巨噬細胞內質網應激
2021-06-17 12:57:34任晶晶彭其勝
中國獸醫學報 2021年5期
關鍵詞:檢測
任晶晶,李 晨,彭其勝
(吉林大學 人獸共患病研究所/教育部人獸共患病研究重點實驗室, 吉林 長春 130062)
布魯菌病(簡稱布病)是由布魯菌侵入機體后引起的傳染-變態反應性人獸共患細菌病,被列為我國法定傳染病中的乙類傳染病之首。布魯菌是典型的胞內寄生菌,其毒力在于細胞內生存和增殖[1]。內質網是細胞內蛋白質合成、加工運輸和分泌的主要場所,對于維持細胞穩態具有重要作用。內質網不僅在機體抵抗胞內感染中發揮重要作用,并且還是多種胞內寄生菌逃避宿主免疫殺傷的理想靶位點[2]。布魯菌的入侵和增殖同樣與內質網有著緊密的聯系[3]。研究發現布魯菌在宿主細胞的內質網內生存與繁殖,能導致內質網的結構重組,產生內質網應激[1]。近年來學者們發現布魯菌感染會誘發內質網應激中的未折疊蛋白反應[4-6]。
布魯菌分泌的效應蛋白可以分為Ⅳ型分泌系統依賴性的和Ⅳ型分泌系統非依賴性的2種。目前國內外研究表明,Ⅳ型分泌系統依賴性效應蛋白在調節布菌感染誘發內質網應激反應中發揮重要調節作用[4-5]。但是,隨著對布魯菌感染機制的深入研究,學者們發現了4種比較特殊的效應蛋白BPE159、BspG、Bsp和BspK,這些分泌到宿主細胞內的效應蛋白不依賴于Ⅳ型分泌系統的存在[5,7],即使布菌的Ⅳ型分泌系統被缺失,它們依然能分泌到宿主細胞的包漿內。然而,關于Ⅳ型分泌系統非依賴效應蛋白調節內質網應激反應的功能目前未知。
通過分析這4種Ⅳ型分泌系統非依賴性蛋白,發現BspI蛋白由布魯菌的一號染色體基因BAB1-1865編碼,含有3個獨特的結構域:信號肽域(1~17 aa)、轉膜域(17~33 aa)及GTPase活化域(98~220 aa)[5]。其中GTPase活化域理論上能夠調節內質網小G蛋白的活性,影響內質網的組裝,暗示著可能參與布魯菌感染引起的內質網應激[8]。因此,本研究選取Ⅳ型分泌系統非依賴效應蛋白BspI作為研究對象,探究其在未折疊蛋白反應中的相關調節功能。
1 材料與方法
1.1 質粒與試劑PGEX-4T-1載體購自吉林安舍科技公司;T4DNA連接酶、限制性核酸內切酶為TaKaRa公司產品;DNA Marker、衣霉素為北京索萊寶公司產品;膠回收純化試劑盒和質粒小提試劑盒為天根公司產品;Anti-rabbit IgG antibody-HRP購自Bioworld公司;Anti-Bip IgG購自Affinity公司;Anti-XBP1 IgG和Anti-his IgG購自Santa公司;細胞因子ELISA檢測試劑盒購自博士德生物公司;反轉錄試劑盒購自莫納公司;細胞RNA提取試劑盒購自天根公司;CCK-8試劑盒購自MCE公司;Glutathione Beads購自天地人和生物有限公司。
1.2 引物的設計與合成以滅活(80℃,2 h)的S19菌液為模板,根據布魯菌S19標準株的BspI基因(NC_007618.1)設計上游和下游引物進行擴增。設計BspI基因引物序列見表1,由吉林庫美生物公司合成。擴增程序按照Premix TaqTM說明書設置(TaKaRa公司Cat.#:RR901)。
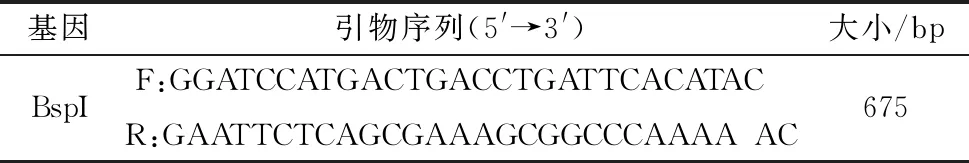
表1 引物序列及擴增片段大小
1.3 GST-BspI融合蛋白的表達、純化及Pull-down試驗首先運用基因克隆方法構建PGEX4T-1-BspI原核表達載體,然后按照文獻[9]進行GST-BspI融合蛋白的表達純化及Pull-down試驗。將SDS-PAGE切膠,送到上海毆易生物公司進行質譜分析。
1.4 BspI蛋白的原核表達、純化及鑒定將目的基因插入pET-28a(+)載體中,構建pET-28a-BspI原核表達載體,經PCR及測序鑒定正確后,轉入大腸桿菌BL21(ED3)感受態細胞,以1.0 mmol/L的IPTG誘導重組蛋白BspI表達。利用AKTA蛋白純化系統純化BspI蛋白。以Anti-his tag IgG為一抗,HPR標記的羊抗鼠IgG為二抗,對純化后的重組蛋白進行Western blot鑒定。
1.5 CCK-8細胞毒性試驗將RAW264.7細胞鋪于96孔板中,分別設置試驗組、對照組以及空白組。試驗組分別加入不同質量濃度的BspⅠ蛋白(10,20,40,80 mg/L),每組設置3個平行孔,置于5% CO2、37℃的細胞培養箱中培養24 h。然后每孔加入10 μL CCK-8溶液,繼續培養1 h后使用酶標儀測定各孔D450 nm值,計算各組細胞存活率。
1.6 BspI蛋白對RAW264.7細胞Bip及IL-6和TNF-α表達影響將RAW264.7細胞鋪于6孔板中,分別將質量濃度為40,60 mg/L的BspI蛋白加入到6孔板中以刺激細胞。設置陽性對照即內質網應激激活劑衣霉素作用組及陰性對照組。24 h后利用Western blot檢測Bip蛋白表達。用雙抗夾心ELISA試劑盒檢測IL-6和TNF-α的表達。
1.7 BspI蛋白對RAW264.7細胞未折疊蛋白反應相關分子mRNA水平的影響為檢測未折疊蛋白反應相關分子IRE1、PERK、ATF6、GAPDH、XBP1和ATF4在mRNA水平的表達量,根據所需要檢測基因在基因庫中mRNA的序列,設計IRE1 (XM_005694366.1)、PERK(XM_005686691.1)、ATF6(AY942654)、GAPDH(XM_005680968.1)、XBP1(XR_3106431)和ATF4(NM_001142518.1)基因的定量引物,引物序列見表2。利用細胞RNA提取試劑盒提取經BspI刺激24 h后的RAW264.7細胞RNA,再利用反轉錄試劑盒將RNA反轉錄為cDNA,進行qPCR檢測。反應程序:95℃ 30 s,95℃ 15 s,60℃ 1 min,共34個循環。

表2 引物序列及擴增片段大小
1.8 BspI蛋白對RAW264.7細胞未折疊蛋白反應相關蛋白表達的影響取BspI刺激24 h后的RAW264.7細胞及陰性對照利用Western blot檢測XBP-1表達情況,同時利用免疫熒光檢測XBP-1的表達。免疫熒光樣品制備步驟:用4%多聚甲醛4℃固定30 min,PBS洗滌后用0.1%曲拉通-100作用10 min。PBS洗2次后用5%BSA室溫固定1 h,以Anti-XBP-1為一抗,1∶100稀釋后4℃過夜。PBS洗2次,二抗1∶500稀釋后室溫避光孵育1 h,最后在熒光顯微鏡下進行觀察。
2 結果
2.1 GST-BspI融合蛋白的表達純化及質譜分析以布魯菌S19株滅活菌液為模板,擴增出BspI基因,結果顯示在675 bp處有單一DNA條帶,與預期大小相符(圖1A)。將構建的PGEX-4T-1-BspI轉化至大腸桿菌BL21中,IPTG誘導表達后純化。SDS-PAGE結果顯示,成功純化出GST-BspI融合蛋白(圖1B)。與RAW264.7細胞Pull-down后可見差異蛋白,進行質譜分析發現BspI可與Bip蛋白結合(圖1C)。
2.2 BspI蛋白的原核表達、純化及鑒定將構建成功的pET28a-BspI載體轉化至大腸桿菌BL21中,用IPTG誘導BspI蛋白表達。SDS-PAGE分析結果顯示,重組BspI蛋白在37℃條件下表達量較16℃條件下表達量多,且大部分形成包涵體,少部分為可溶蛋白(圖2A)。利用AKTA蛋白純化系統對可溶蛋白進行純化,SDS-PAGE分析后結果顯示,400 mmol/L咪唑洗脫效果最優(圖2B)。Western blot分析證明,重組BspI蛋白可以被His-tag抗體特異性識別(圖2C)。

A.BspI蛋白可溶性分析(M1.蛋白質相對分子質量標準,1.pET28a(+)空載體誘導產物,2.37℃誘導pET28a(+)-BspI全菌,3.37℃誘導pET28apET28a(+)-BspI超聲上清,4.37℃誘導pET28apET28a(+)-BspI超聲沉淀,5.16℃誘導pET28apET28a(+)-BspI全菌,6.16℃誘導pET28apET28a(+)-BspI超聲上清,7.16℃誘導pET28apET28a(+)-BspI超聲沉淀);B.不同咪唑濃度洗脫液SDS-PAGE分析(M2.蛋白質相對分子質量標準,8.20 mmol/L咪唑洗脫液,9.50 mmol/L咪唑洗脫液,10.200 mmol/L咪唑洗脫液,11.400 mmol/L咪唑洗脫液,12.500 mmol/L咪唑洗脫液);C.BspI重組蛋白Western blot鑒定(13.誘導空載體菌液,14.37℃誘導菌液)
2.3 BspI對RAW264.7細胞存活率的影響CCK-8試驗結果(圖3)顯示,BspI質量濃度為80 mg/L時,細胞存活率顯著性下降(P<0.001),而質量濃度為40 mg/L時,細胞存活率與對照組相比并沒有明顯變化。因此后續采用質量濃度為40,60 mg/L 的BspI刺激細胞,檢測未折疊蛋白反應發生,明確BspI刺激質量濃度。
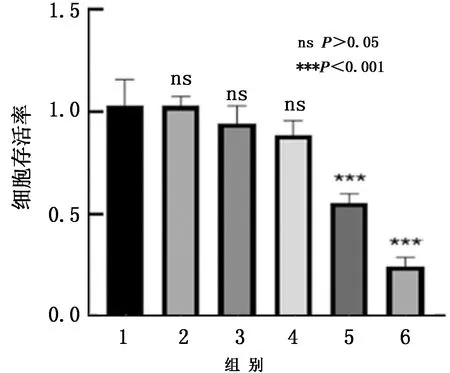
1.對照組;2.10 mg/L BspI蛋白;3.20 mg/L BspI蛋白;4.40 mg/L BspI蛋白;5.80 mg/L BspI蛋白;6.160 mg/L BspI蛋白
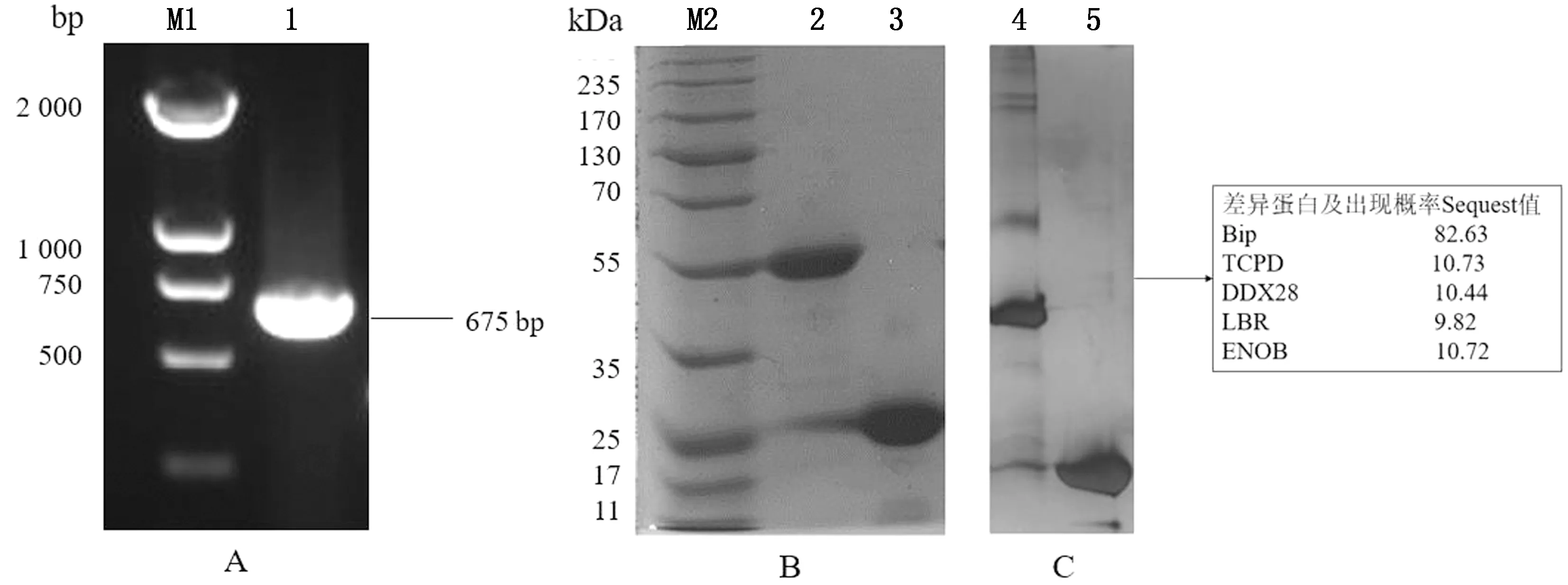
A.BspI基因PCR擴增(M1.DL2000 DNA Marker,1.BspI基因的PCR產物);B.GST-BspI融合蛋白的純化(M2.蛋白質相對分子質量標準,2.純化后的GST-BspI融合蛋白,3.純化后的GST蛋白);C.質譜差異蛋白分析(4.GST-BspI融合蛋白Pull-down結果,5.GST蛋白Pull-down結果)
圖1 GST-BspI表達載體鑒定、融合蛋白的表達純化和質譜差異蛋白分析
2.4 BspI蛋白對RAW264.7細胞未折疊蛋白反應標志性分子Bip及相關細胞因子表達影響收集衣霉素處理的陽性對照組、PBS處理的陰性對照組及BspI蛋白刺激組的細胞,裂解后進行Western blot檢測。結果如圖4A所示,質量濃度為60 mg/L的BspI蛋白刺激RAW264.7細胞24 h后,Bip蛋白表達量顯著升高,引起的未折疊蛋白反應更為顯著。由于TNF-α和IL-6與未折疊蛋白反應有關,因此對TNF-α和IL-6進行ELISA檢測,結果顯示(圖4B,C),RAW264.7細胞TNF-α和IL-6的表達顯著提高(P<0.001,P<0.05)。
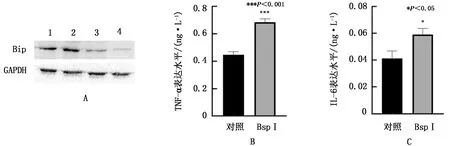
A.不同質量濃度BspI重組蛋白刺激RAW264.7細胞Bip表達量鑒定(1.衣霉素陽性對照組,2.60 mg/L BspI作用組,3.40 mg/L BspI作用組,4.PBS陰性對照組);B.BspI對RAW264.7細胞TNF-α表達的影響;C.BspI對RAW264.7細胞IL-6表達的影響
2.5 BspI蛋白對RAW264.7細胞未折疊蛋白反應相關分子轉錄水平影響上述結果表明BspI可以引起RAW264.7細胞的未折疊蛋白反應。為明確相關通路,利用實時熒光定量PCR檢測BspI對參與未折疊蛋白反應的3條信號通路相關分子mRNA轉錄的影響。結果表明,BspI蛋白刺激RAW264.7細胞24 h后,IRE1與XBP-1的mRNA表達量較對照組有顯著升高(圖5C,E;P<0.01,P<0.05),而PERK、ATF6與ATF4的mRNA表達量較對照組無明顯變化(圖5A,B,D)。
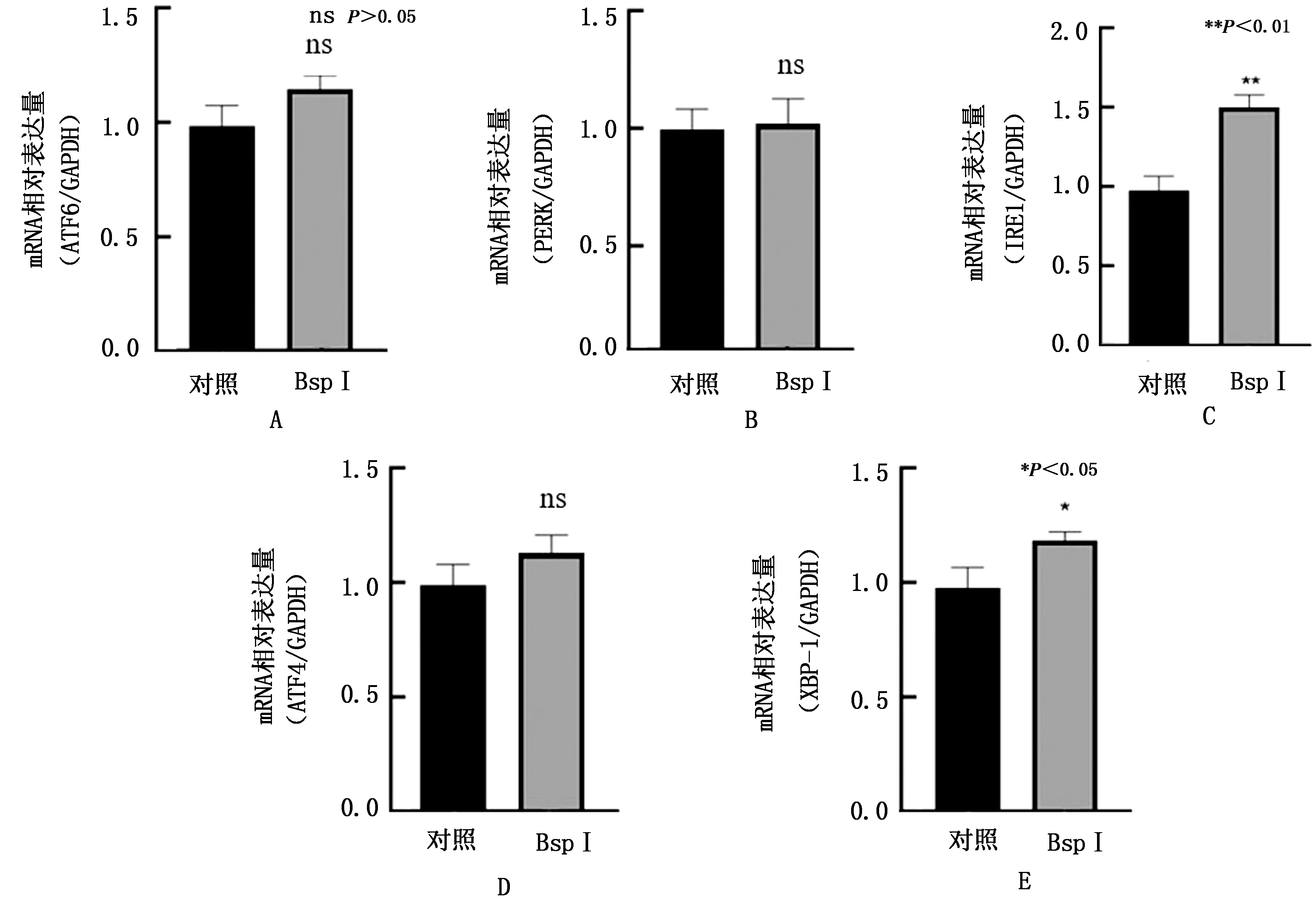
A.ATF6;B.PERK;C.IRE1;D.ATF4;E.XBP-1
2.6 BspI蛋白對RAW264.7細胞XBP-1蛋白表達的影響分別利用Western blot與免疫熒光檢測試驗組與對照組的XBP-1蛋白表達情況,結果顯示(圖6),試驗組的XBP-1蛋白表達水平顯著升高,與對照組相比有極顯著性差異(P<0.001)。
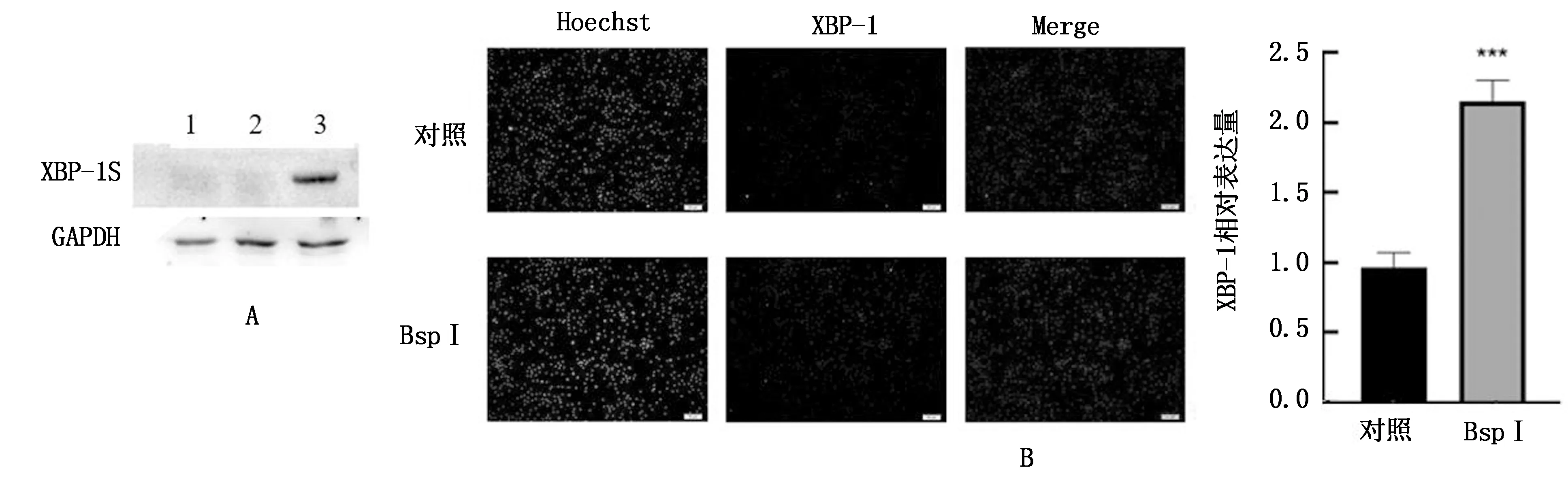
A.XBP-1S在不同處理組的表達(1.TM陽性對照組,2.PBS陰性對照組,3.BspI處理組);B.免疫熒光檢測XBP-1在RAW264.7細胞表達
3 討論
布魯菌感染宿主細胞后,通過細胞內吞作用進入細胞質中,形成早期布魯菌菌泡,歷經中期布魯菌菌泡,最后與細胞內質網發生融合,形成復制型布魯菌菌泡。從此,布魯菌在細胞內開始增殖,顯示其毒力[10-12]。目前已知布魯菌在與細胞內質網互作時,能誘導細胞內質網應激,通過未折疊蛋白反應等實現內質網功能穩定,促進布魯菌的胞內生存[13]。用TUDCA(一種未折疊蛋白反應的抑制劑)處理宿主細胞能顯著抑制布魯菌在細胞內的增殖[6,14]。可見未折疊蛋白反應在布魯菌的胞內生長中發揮重要作用。PERK、IRE1和ATF6是未折疊蛋白反應的3條信號通路的重要分子。當未折疊蛋白反應未發生時, Bip能結合這3個分子的內質網內端,使它們處于非活化狀態。當未折疊蛋白反應發生時,Bip表達增加,Bip與未折疊蛋白結合,解除對這3個分子的抑制作用,激活UPR反應的3條信號通路,恢復細胞內環境穩態。由此也可見,Bip是未折疊蛋白反應的關鍵性調控分子[13]。
本研究發現布魯菌BspI蛋白能夠與Bip結合,因此推測布魯菌BspI蛋白能夠引起宿主細胞的未折疊蛋白反應。為此,首先表達并純化BspI蛋白,確定BspI刺激所需質量濃度后通過Western blot檢測發現未折疊蛋白反應標志分子Bip蛋白表達量顯著增加。同時,通過實時熒光定量PCR、Western blot和免疫熒光試驗發現巨噬細胞在受到BspI蛋白刺激后,巨噬細胞中的XBP-1S表達量增加。XBP-1由無活性的XBP-1U轉化為有活性的XBP-1S是未折疊蛋白反應發生的重要標志之一。表明BspI激活了巨噬細胞未折疊蛋白反應,從而證明布魯菌BspI蛋白在巨噬細胞內質網應激調控中發揮重要作用
布魯菌感染導致的巨噬細胞內質網應激能誘導炎癥因子,如TNF-α和IL-6的產生[15]。在本研究中利用ELISA試劑盒檢測巨噬細胞受到BspI刺激后,細胞培養上清液中TNF-α與IL-6分泌量情況。結果發現BspI蛋白刺激可以顯著引起炎性因子TNF-α與IL-6的分泌,這從另一個角度證明BspI能誘導巨噬細胞內質網應激的發生。
綜上所述,本研究證明布魯菌BspI蛋白可以引起巨噬細胞內質網應激反應,且主要是通過未折疊蛋白反應中的IRE1通路。同時還提出了Ⅳ型分泌系統非依賴性效應蛋白在布魯菌感染導致的內質網應激反應中發揮重要調控作用。本研究發現Ⅳ型分泌系統非依賴性效應蛋白也可以參與布魯菌感染導致的宿主細胞內質網應激的調控,豐富了布魯菌感染的機制研究,為未來進一步研究內質網應激反應提供新思路。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
