棉花花青素能量耗散過程維持光合機構穩定性分析
2021-06-07 06:01:16李霞飛李自良賈夢夢陳云瑞張旺鋒張亞黎
新疆農業科學 2021年5期
李霞飛,李自良,賈夢夢,陳云瑞,向 導,張旺鋒,張亞黎
(1.石河子大學農學院/新疆生產建設兵團綠洲生態農業重點實驗室,新疆石河子 832003;2.新疆烏蘭烏蘇農業氣象試驗站/烏蘭烏蘇國家綜合氣象觀測專項試驗外場,新疆烏蘭烏蘇 832199)
0 引 言
【研究意義】花青素主要通過過濾、衰減和反射高強度的藍紫光從而調節對光能的吸收[1],陳靜等[2]研究低溫弱光下光化學能力降低導致過量激發能的積累,而花青素的存在能夠維持一定的光合能力。花青素的存在能夠減少PSII中的過剩激發能,有效保護PSII的光合活性[3-4];薛占軍等[5]研究花青素通過吸收400~600 nm的可見光,有效保護PSI的活性。張開明等[6]研究發現,含有花青素的紅葉光合速率較低,主要是由于花青素限制了葉綠體對光能的吸收,影響光反應中心的開放從而影響碳同化過程,花青素與植物光合作用有密切聯系[7]。分析含有花青素的棉花光合參數的變化,對研究花青素作為光保護機制的研究有重要意義。【前人研究進展】植物生育進程中常出現各種逆境脅迫,將導致光合器官吸收的光能不能全部用來進行光化學反應,過剩的光能會產生大量活性氧自由基,破壞植物光合機構,影響光合作用的穩定運行[8]。為了防止光損傷的發生,植物激發了各種光保護機制來減少過剩激發能的積累,包括葉片運動[9]、以無害熱量形式散失的熱耗散[10]、通過光化學途徑產生能量物質促進D1蛋白的合成,修復光反應中心[11-12],以及圍繞PSI的環式電子傳遞[13-14]。花青素也可以作為一種光保護機制維持光合機構的穩定運行[15]。依賴葉黃素循環的熱耗散[16-18]是保護光合機構免受損傷的重要的光保護機制,類黃酮類物質在長期的光保護過程中比葉黃素循環更為重要[19],但也有研究表明花青素并不具有光保護作用[20]。【本研究切入點】花青素可以作為一種光保護機制保護光合機構免受損傷。有關花青素和其他光保護機制環式電子流和葉黃素循環的熱耗散之間關系的研究較少。以不同花青素含量的棉花為材料,研究花青素含量的差異對其光合性能的影響以及為維持光合機構運行所啟動的光保護機制。【擬解決的關鍵問題】分析花青素含量差異與光保護機制的關系,為棉花的高光效育種提供理論依據。
1 材料與方法
1.1 材 料
試驗于2018年在新疆中國氣象站烏蘭烏蘇生態與農業氣象站(44°17’N,86°03’E)進行。供試材料由國家棉花種質資源中期庫(National Medium-term Gene Bank of Cotton in China)和國家棉花種質資源平臺(National cotton germplasm resources platform)提供。圖1,表1

圖1 供試材料

表1 供試材料編號及名稱
1.2 方 法
1.2.1 試驗設計
小區面積4.5 m2,隨機區組排列,重復3次。4月22日布滴灌帶,鋪膜后人工點播,2 d后進行少量灌溉,保證出苗。寬窄行種植,株距12 cm,密度為22×104株/hm2,其他管理措施同一般大田管理。
待棉花進入盛花期,葉片顏色均勻一致,取主莖上倒數第1片完全展開葉,每個材料取3次重復進行試驗。
1.2.2 指標測定
1.2.2.1 葉綠素
將取回的棉花葉片用打孔器(孔徑5 mm)打成小圓片4個,放入 5 mL離心管,用約4 mL 80%的丙酮溶液浸提,置于完全遮光處浸提,定時振蕩至圓片完全褪綠。取其上清液用分光光度計UV-2041 型分光光度計(日本島津)比色,葉綠素含量的計算公式參考Lichtenthaler[21]。
1.2.2.2 花青素
選顏色均勻的棉花葉片,在葉脈兩側用5 mm打孔器取15個葉圓片,用15 mL 1%鹽酸甲醇溶液提取,在黑暗環境中浸提24~48 h,25 mL容量瓶定容,1%鹽酸甲醇溶液為空白對照。在波長530和652 nm處測其吸光度。花青素含量花青素(Ant)=A550-0.25A652[22]。
1.2.2.3 光合參數
Li-6400便攜式光合測定系統(Li-COR,USA)測定光合氣體參數凈光合速率(Pn)。測定時,設置光強 2 000 μmol/(m2·s)、CO2濃度(400±2) μmol/(m2·s)、氣體流量300 μmol/s,葉片溫度(30±2)℃,參比室的水分(20±2) μmol/(m2·s)。將棉花葉片左上部加入葉室,待數據穩定后,記錄數據。
1.2.2.4 吸光系數
用Imaging-PAM熒光成像儀(Walz,德國)測定吸光系數。首先校準儀器,然后將被測材料放置到成像區域,用樣品架上的細線固定材料,在材料中上部葉脈兩側選擇5~6個區域,測定吸光系數(Abs)。
1.2.2.5 NDH活性
利用Dual-PAM-100葉綠素熒光儀(Walz,德國)測量NDH活性和P700暗還原速率。將取回的棉花材料放置在室溫條件下進行0.5~1 h黑暗適應,打開光化光AL(1 828 μmol/(m2·s))進行熒光誘導,待熒光信號趨于平緩狀態(大概4 min左右),關掉光化光熒光信號先下降后立即上升,熒光信號立即上升速率能夠反映NDH介導的環式電子活性[23]。
1.2.2.6 P515吸收變化
單周轉飽和閃光誘導活體棉花葉片的P515快速馳豫動力學可以反映類囊體膜的完整性和ATP合成酶活性[24]。經過1 h暗適應后,P515吸收信號先快速上升,緩慢上升,達到峰值后,緩慢下降。先打開光化光(1 800 μmol/(m2·s))照光10 min,后暗適應4 min,P515吸收信號先快速上升后快速下降,緩慢上升相消失。暗適應后P515吸收信號緩慢下降,膜的完整性,照光后P515吸收信號快速下降的初始斜率可以用來表示ATP酶活性[25]。
利用Dual-PAM-100(Walz,Germany)的新模塊P515/535 檢測雙波長550~515差示吸收的信號變化。將供試材料充分暗適應(至少1 h),測定的515信號能夠反映類囊體膜的完整性[24],然后打開光化光(1 828 μmol/(m2·s))誘導棉花葉片10 min,關掉光化光黑暗處理4 min,再測定P515信號吸收變化能夠反映ATP合成酶活性[25]。
測定550~515 nm信號的暗-光-暗誘導瞬變,取充分暗適應幾個小時的棉花材料,打開光化光AL(1 828 μmol/(m2·s))誘導10 min后515信號趨于穩定(玉米黃質合成的測定),然后關掉光化光AL,515信號先快速下降隨后上升逐漸趨于穩定(跨類囊體膜質子梯度的測定)[26-27]。
1.3 數據處理
采用Microsoft Excel 2019進行數據處理,運用Rstudio軟件進行方差分析及作圖。試驗處理間的多重比較采用“Agricolae”程序包進行最小顯著差法(Least-significantdifference,LSD)分析,顯著水平為0.05,并用英文小寫字母標記。圖表中的數據均為平均值±標準誤。
2 結果與分析
2.1 不同棉花花青素含量的差異
研究表明,綠葉棉花無花青素的存在,而紅葉棉花有明顯花青素存在,隨顏色加深,花青素含量增多。低花青素含量的棉花Chla、Chlb、ChlT均較高花青含量的棉花低,而無花青素的棉花 Chla、Chlb、ChlT和高花青素含量棉花幾乎無差異,低花青素棉花類胡蘿卜素(Car)較高花青素棉花低,含花青素的棉花較無花青素的棉花低。圖2
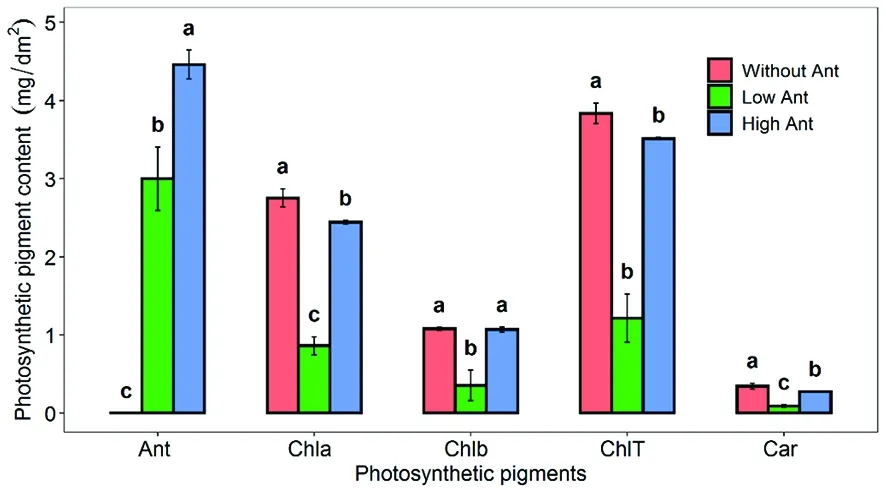
圖2 不同棉花花青素含量和葉綠素含量的差異
2.2 不同花青素含量的棉花光合性能的差異
研究表明,含花青素和不含花青素的棉花Fv/Fm均無差異,各棉花可能均相應啟動了光保護機制。含花青素的棉花Abs較不含花青素的棉花高,含花青素的棉花吸光系數(Abs)為0.87明顯高于較不含花青素的棉花0.83,低花青素棉花Abs 0.86和高花青素棉花0.87無差異;隨花青素含量增加,凈光合速率(Pn)從31.19到31.43無差異,而無花青素的棉花Pn 34.25顯著高于含花青素的棉花31.19。表2
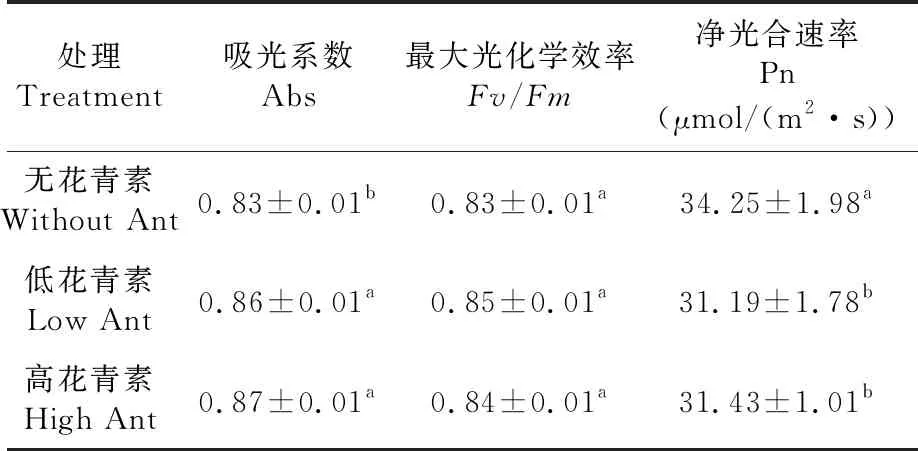
表2 不同花青素含量的棉花氣體交換參數變化
2.3 不同花青素含量的棉花NDH介導的環式電子活性的差異
研究表明,隨花青素含量增加,熒光信號的快速上升速率顯著增強,而花青素含量較低的棉花和無花青素的棉花熒光信號的快速上升無明顯差異。花青素含量越高NDH介導的環式電子活性越高,高花青素的棉花伴隨較強的NDH介導的環式電子流。圖3

A:葉綠素熒光動力學曲線;B:作用光關閉后葉綠素熒光的瞬時上升
2.4 不同花青素含量的棉花ATP合成酶活性的差異
研究表明,在Before AL條件下高花青素含量的棉花和無花青素的棉花515信號緩慢上升幾乎無差異,類囊體的完整性都相對較好;而After AL條件下高花青素含量的棉花515信號快速下降的初始斜率顯著高于無花青素的棉花。高花青素含量的棉花較無花青素的棉花有較高的ATP合成酶活性,能夠合成更多的ATP,而低花青素含量的棉花和無花青素的棉花有相同的趨勢。圖4
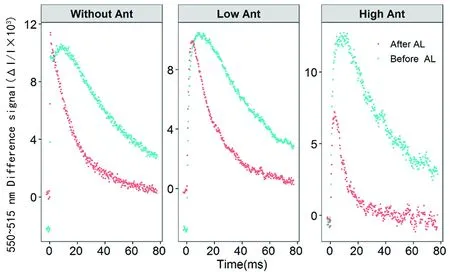
圖4 花青素含量的差異對單周轉飽和閃光誘導P515的吸收變化
2.5 不同花青素含量的棉花葉黃素循環的熱耗散的差異
研究表明,在515信號暗-光誘導下,無花青素的棉花515信號顯著較含花青素的棉花強,而低花青素含量的棉花較高花青素含量的棉花強,在含花青素的棉花中玉米黃質的合成速率顯著較無花青素的棉花弱,隨花青素含量的增多,玉米黃質的合成速率變慢,花青素的存在可能抑制了玉米黃質的合成,并且隨花青素含量的增多,抑制作用越強。圖5
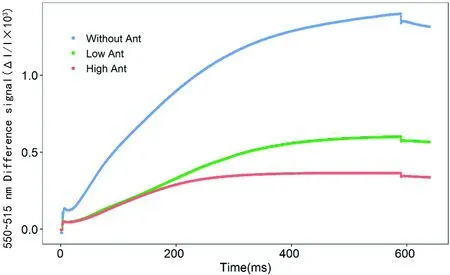
圖5 花青素含量的差異下550~515 nm信號的暗-光-暗誘導瞬變的影響誘導瞬變
3 討 論
3.1 花青素含量增多伴隨葉片光合能力的減弱
花青素可通過過濾、衰減和反射高強度的藍紫光從而來減少光能的吸收[15、27],可以調節葉綠素對光量子的吸收[1]。而光合色素在植物光合作用中主要起對吸收、傳遞和轉化光能的作用,其中Chlb具有捕獲和吸收光能的作用,而Chla能夠執行能量轉換,類胡蘿卜素不僅可以作為輔助色素捕獲光能,也可以猝滅活性氧防止光合器官受損[28]。高花青素含量的棉花Chla、Chlb、ChlT、Car均低花青素含量的高,吸光系數之間無差異,而相對無花青素的棉花而言,高花青素含量的棉花Chla、Chlb、ChlT較低,吸光系數較高,Car較低。花青素含量越低,吸收、轉換光能的能力較弱,猝滅活性氧的能力也較弱,花青素含量的多少能夠影響植物對光能的吸收以及猝滅活性氧的能力。
分析凈光合速率,低花青素棉花的Pn與較高花青素棉花無差異,相對無花青素的棉花,有花青素的棉花Pn較弱,有花青素的存在并且隨著花青素含量逐漸增多,光合性能越弱,可能含有花青素的棉花吸收的光能被花青素吸收、過濾使得轉換為光化學的能量減弱,使得光合能力減弱。
3.2 隨花青素含量升高NDH介導的環式電子流及其調控能力逐漸增強
環式電子傳遞是植物體內重要的光合電子傳遞途徑[13、29],研究中,很明顯高花青素含量的棉花中NDH介導的環式電子活性顯著高于無花青素的棉花,ATP合成酶活性也較無花青素的棉花強;同時隨花青素含量的增加,Y(ND)逐漸增強,Y(NA)逐漸減弱,較強的環式電子能夠耗散受體側的電子,減少氧化態的P700,保護PSI免受光損傷,有效保護PSI的活性。進一步分析高花青素含量的棉花ATP合成酶活性較低花青素含量的棉花強,并且高花青素含量的棉花能夠較好的保護PSII的活性。隨花青素含量的增加,棉花光合能力減弱,而吸光能力增強,將面臨較多的過剩激發能,花青素含量的增加伴隨環式電子流及其調控能力的增強,有效保護了PSI和PSII的活性。
3.3 花青素能夠起到耗散過剩光能保護光合機構的作用
P515信號的暗-光-暗誘導瞬變曲線能夠反映玉米黃質的合成速率和類囊體腔內H+的積累程度[26]。515光-暗誘導下,515信號最低點和穩態點之間的差值能夠反映ΔpH[27],無花青素的棉花ΔpH明顯較含有花青素的棉花高,不含花青素條件下類囊體腔內的H+濃度顯著較高。
分布在葉片上表皮的花青素能夠阻擋強光[30],Gould等[31]發現含花青素的葉片在受到光損傷后能夠快速恢復;Neill和Gould的工作證實,花青素可以耗散過剩激發能減少活性氧的產生[32]。Hughes等[33]研究證實,紅葉樹種的葉黃素循環過程較弱,更多的依賴花青素的光保護作用。
研究中,含有花青素的棉花玉米黃質的合成速率顯著低于無花青素的棉花,可能是紫黃質轉化為玉米黃質的過程受到了影響,跨類囊體膜質子梯度(ΔpH)也顯著低于無花青素的棉花,也就是說質子梯度驅動的葉黃素循環的熱耗散在含有花青素的棉花中較弱,可能含有花青素的棉花主要依賴花青素的光破壞防御功能保護其光合機構免受光抑制。隨花青素含量的增加,玉米黃質的合成速率減弱,花青素越多,葉黃素循環的熱耗散越弱。相反對于無花青素的棉花來說,主要依賴葉黃素循環的熱耗散維持PSII的穩定。
綠葉棉花不含花青素,主要依賴環式電子途徑建立的ΔpH驅動葉黃素循環的熱耗散過程保護光合機構免受光損傷;而含有花青素的棉花主要依賴環式電子建立的質子梯度驅動ATP合成酶合成ATP,合成D1蛋白修復PSII,減少氧自由基的傷害,另一方面花青素本身具有一定的光破壞防御功能替代了葉黃素循環的過程耗散過剩激發能,保護光合機構。
4 結 論
隨花青素含量的增加,凈光合速率從34.25減弱至31.43,而吸光系數由0.83增加至0.87,因此,將面臨較多的過剩激發能。關關熒信號的快速上升的斜率也逐漸增強,并且After AL條件下515信號快速下降的初始斜率也顯著升高,光-暗反應條件下515信號葉黃素循環的熱耗散能力逐漸減弱;無花青素的棉花515信號葉黃素循環的熱耗散顯著較含花青素的棉花強。含花青素的棉花可以通過花青素的耗散過程維持光合機構的穩定,而無花青素的棉花主要通過依賴葉黃素循環的熱耗散耗散過剩光能保護光合機構免受損傷。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年3期)2019-02-01 06:12:26
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
