午潮山常綠次生闊葉林主要木本植物功能群及其演替特征
2021-06-04 03:27:38吳初平袁位高朱錦茹黃玉潔焦潔潔臧潤國
生態學報 2021年8期
金 超,吳初平,丁 易,袁位高,朱錦茹,黃玉潔,焦潔潔,臧潤國,江 波,*
1 浙江農林大學 林業與生物技術學院, 杭州 311300 2 浙江省林業科學技術研究院, 杭州 310023 3 中國林業科學研究院森林生態環境與保護研究所,國家林業和草原局森林生態環境重點實驗室, 北京 100091 4 南京林業大學南方現代林業協同創新中心, 南京 210037
次生林是目前大多數森林所處的演替狀態[1],了解其群落組成和物種關系對于保護森林多樣性和碳匯具有重要意義[2- 3]。研究群落次生演替過程中的物種組成和生態系統功能可以提供促進群落正向演替的信息[4]。其中,分析不同生活史策略的物種相互關系及其與生態系統功能的聯系可以幫助理解和預測群落演替過程[5]。探索次生林不同演替階段的物種間關系為管理和修復森林資源提供極大的便利。
評價并劃分群落中具有相似功能的物種可以幫助量化物種對環境的響應程度和理解群落構建的過程[6]。根據物種演替地位和最大潛在高度將其劃分功能群是被廣泛接受的方法[7],而數量分類通過計算植物功能性狀相似性劃分功能群,為本研究提供了新視角[8]。植物功能性狀能反映其生態策略[9],其中,葉經濟譜[10]和木材經濟譜[11]能較好地描述物種性能的功能性狀。比如,相較于檫木(Sassafrastzumu)、麻櫟(Quercusacutissima)等喜光的先鋒樹種,青岡(Cyclobalanopsisglauca)等耐陰樹種在弱光照下的壽命更長且生長速度緩慢,其功能性狀表現為低比葉面積和高木材密度[12- 14]。根據實驗目的選取植物功能性狀將物種劃分到對應的功能群,為研究復雜的生態系統過程建立基礎[15- 16]。有研究利用功能群劃分來研究物種組成復雜的熱帶山地雨林[17],此外,還有分析不同生境條件下植物功能群和物種性狀的響應,揭示了土壤異質性的重要性[18]。
功能群內和功能群之間的生態位和物種聯結可以反映物種對生境資源的競爭關系,進而加強對群落結構、功能及演替的理解。生態位指物種和環境之間相互影響和相互作用的關系[19],它表達了群落中物種對資源的競爭程度,競爭者間的生態位差異代表良好的共存關系[20],而種間聯結反映物種的空間分布和功能依賴關系[21]。其概念廣泛應用于研究群落組成及指導植被的恢復與重建[22- 24],但由于缺乏植物功能的機制基礎,直接比較群落物種的生態位和種間關系僅能得出片面的結論。Fry等指出通過植物性狀劃分功能群比簡單的草本、灌木和喬木劃分更能體現生物多樣性和生態系統功能的關系[25]。因此,利用功能群探索物種關系能為研究森林生態系統提供重要的實際應用價值。
午潮山位于杭州西郊,曾經過度的森林砍伐和不合理的土地利用導致大部分原始植被被破壞,經過長期的改造和恢復,形成以木荷(Schimasuperba)、青岡、苦櫧(Castanopsissclerophylla)為建群種的次生林[26]。由于受到早期種植的針葉樹種影響,午潮山森林的林分結構和演替進度在一定程度上被影響。鑒于此,本研究選取午潮山常綠次生闊葉林70個樣地中的主要物種進行功能群劃分,并評估不同功能群間的物種關系。旨在回答以下問題:1)午潮山森林群落功能群內和功能群間的植物生態位和種間關系如何?其處于什么演替階段?2)如何根據功能群和種間關系的相關結論對午潮山常綠次生闊葉林進行生態系統修復以及促進群落演替?
1 研究方法
1.1 研究區概況及樣地調查
本研究在浙江午潮山國家森林公園林區內進行(33°41′ N, 120°00′ E),午潮山林區為典型的亞熱帶常綠次生闊葉林。午潮山平均海拔264 m,最高494.7 m,森林覆蓋率達93%,土壤類型為紅壤。地處亞熱帶氣候類型,年平均溫度為16.1℃,最冷一月的月平均溫度為3.6℃。全年的降雨量為1400.7 mm,年無霜期為240 d,年平均相對濕度為76%,年平均日照時數為1800—2100 h。午潮山原始天然林植被破壞后人為種植大量經濟樹種:濕地松(Pinuselliottii)和杉木(Cunninghamialanceolata),對森林進行保護和恢復后逐漸演替到目前的常綠次生闊葉林。目前,研究區森林群落的喬木層以木荷、青岡、杉木和麻櫟為優勢樹種,灌木層的主要樹種包括毛柄連蕊茶(Camelliafraterna)、薄葉山礬(Symplocosanomala)、馬銀花(Rhododendronovatum)、窄基紅褐柃(Euryarubiginosavar.attenuata)和檵木(Loropetalumchinense)。
分別于2018年4月和2019年10月參照熱帶森林科學研究中心(Center For Tropical Forest Science, CTFS)的原則[27]在午潮山林區的北坡和南坡建立不重疊的森林樣地。在整個景觀中用全站儀設置了70個0.04 hm2(20 m×20 m)的樣地,每個樣地繼續劃分成16個5 m×5 m的小樣方。對樣方內1.3 m位置處胸徑(DBH)≥1 cm的存活木本樹種進行標定并掛牌,記錄樹種名稱、胸徑、樹高和空間坐標,同時記錄每個樣方的海拔、經緯度、坡度、坡向等生境特征。
1.2 重要值和生態位特征
重要值可以通過參數量化物種在群落中地位和作用[28]。生態位寬度能衡量物種利用資源的范圍,可使用Levins指數(BL)和Shannon指數(BS)來表示,而Pianka生態位重疊指數(Oik)表示生態位重疊程度,其數值范圍從0(無重疊)到1(完全共享資源)。
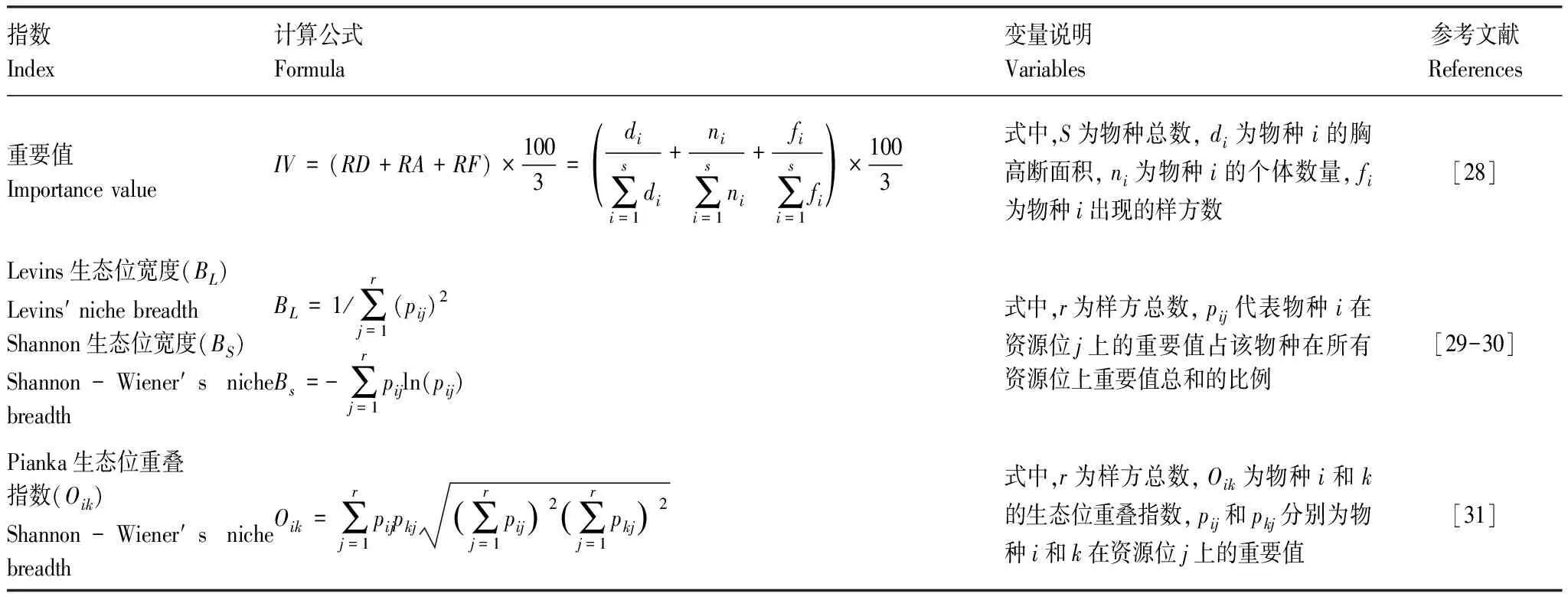
表1 重要值和生態位指數計算公式
1.3 功能群劃分性狀的選擇
計算70個樣地中物種的重要值,將排序前45(重要值占全部物種95%)的木本植物作為劃分功能群的主要物種。演替早期和后期的物種在習性和生態策略有明顯的區別,擁有較高資源獲取率(如喜陽、生長速度快、種子小且多、木材密度低等)的物種在資源豐富的環境占據主導地位,但物種會傾向于發展對極端環境更有忍耐能力和能保存資源的性狀[5]。本文使用與植物生理和形態相關的功能性狀劃分功能群[7, 9- 11],分為分類性狀和連續性狀。分類性狀包括生長型、喜光性和壽命;連續性狀包括葉片氮含量(LNC, mg/g)、葉片磷含量(LPC, mg/g)、葉片鉀含量(LKC, mg/g)、比葉面積(SLA, cm2/g)、木材密度(WD, g/cm3)、葉片干物質含量(LDMC, g/g)、葉片葉綠素含量(ChL, SPAD)、最大潛在樹高(Hmax, m)。生長型通過物種的形態和生長特性劃分為喬木、小喬木或灌木和灌木;物種喜光性則基于亞熱帶常綠闊葉林植被演替階段不同物種的分布特征從喜光到耐陰分為5個等級;物種壽命根據物種早期演替特征利用二分法分為長壽命和短壽命。這些分類變量均反映了物種對環境因子的要求和響應,以及物種在生態系統中的作用和地位。分類變量的數據主要來源于中國植物志網絡版(http://www.iplant.cn/frps),部分缺失的則通過樣地勘測和咨詢植物學教授。
物種的連續性狀數據主要通過實驗獲取。按照國際植物性狀選取標準,選取胸徑10 cm以上的目標物種10株(個體數少的除外)[32],在每株個體的中冠位置(高度≤5 m)采取5片完整、健康的成熟葉片,同時取5—10根枝條(1 cm≤DBH≤2 cm)。使用電子天平、葉面積儀(LI-COR 3100C)和葉綠素儀(SPAD 502 Plus Meter, Konica Minolta, Japan)分別測量葉片鮮重、葉面積和葉綠素含量。最后,將葉片置于65℃的烘箱內烘干至恒重后進行葉片干重測定,隨后對葉片進行葉片氮、磷、鉀含量的測定[33]。用排水法測定去皮后的植物枝條體積,然后將其置于103℃的烘箱內烘干,恒重后稱重。枝條密度利用枝條干重與體積之比計算得出,為了減少樣地內樹種的破壞,使用物種的枝條密度代替木材密度。每個物種的最大潛在樹高信息參考中國植物志網絡版(http://www.iplant.cn/frps)。
1.4 總體聯結性檢驗
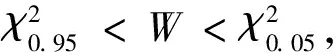

表2 物種聯結性指數計算公式
1.5 種間聯結性分析
1.5.1χ2檢驗
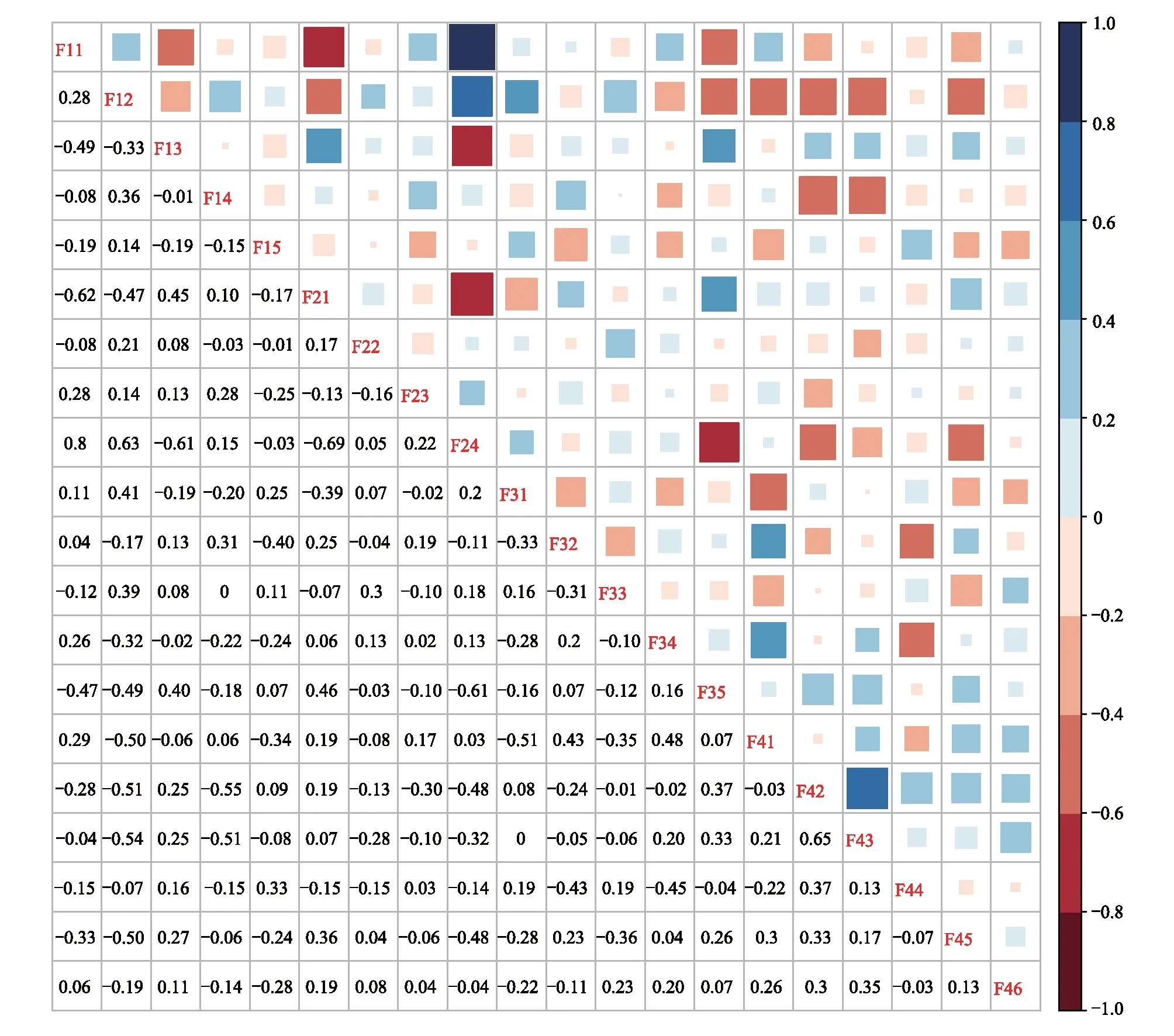
χ2統計量通過使用一個基于2×2列聯表來對種間聯結性進行定性分析[23],由于本實驗為非連續取樣,因此采用Yates的連續校正公式來矯正χ2統計量。若ad-bc>0,種間聯結為正聯結,反之為負聯結。當χ2<3.841,表明物種間是獨立且聯結不顯著(P>0.05);若3.841≤χ2<6.635,說明物種間聯結顯著(0.01 1.5.2Dice值(DI) χ2檢驗僅提供了種間聯結的定性結果,但是不能量化物種對的聯結程度[19],而DI值可以確定其聯結程度。DI的范圍是(0,1),若DI=0,則無聯結,表明物種間完全獨立且幾乎不會同時出現。相反,DI越接近1,物種對就越有可能同時出現。 1.5.3Spearman秩相關系數(rs) χ2檢驗和DI值使用二元數據判斷物種聯結性,因此不可避免的會忽略一些要素,如,物種多度[35]。Spearman秩相關檢驗可以對物種間的線形關系進行定量分析,且包含物種聯結和物種協方差的信息[36],分析種對同時出現的可能性。其值域為[-1,1],正值為正相關,負值為負相關。 本研究中,數據分析在R3.6.0[38]中完成。功能群的劃分使用基于11個功能性狀的聚類分析,采用FD包[39]中的dbFD函數,其優勢是可處理不同類型的功能性狀。由于存在部分數據缺失,分析過程采用Gower相異性矩陣[40- 41],分析方式采用k-均值聚類(k-means partitioning),聚類的組數采用方差比準則(Calinski-Harabasz criterion, VRC)[42],通過比較組間和組內平方和誤差的比值判斷得出,比值越大,聚類效果越好。不同功能群的功能性狀差異用單因素方差分析檢驗,顯著差異使用Tukey檢驗(Tukey Honestly Significant Difference tests,P=0.05)。生態位寬度、生態位重疊程度使用R包“spaa”[43]計算,可視化在“corrplot”包[44]中完成。 本次調查的70個樣地中,共有木本植物13596株119種,分屬40個科76個屬。午潮山林區重要值排序前45的木本植物根據性狀矩陣通過k-均值聚類分析,并結合方差比準則聚類為4個功能群。PFG1由6個物種組成,主要由麻櫟和濕地松等早期喜陽先鋒樹種組成;PFG2由11個物種組成,主要是賽山梅(Styraxconfusus)、小葉石楠(Photiniaparvifolia)和白檀(Symplocospaniculata)等中期中性樹種;PFG3包含15個物種,主要為木荷、青岡和杉木等中后期樹種組成;PFG4則由13種后期耐陰樹種組成,主要為毛柄連蕊茶、薄葉山礬和馬銀花等(表3)。4個功能群之間的連續功能性狀差異顯著(圖1),從物種組成和性狀來看,PFG1到PFG4代表了由演替初期過渡到演替后期的物種。具體來說,物種性狀由喜光、短壽命、低葉片營養物質和低木材密度過渡到耐陰、長壽命、高葉片營養物質和高木材密度。 表3 功能群劃分 圖1 午潮山4個功能群性狀差異(平均值±標準差)Fig.1 The difference of plant functional traits in four functional groups in Wuchao Mountain (mean±SD)P值代表組間差異顯著性水平,不同的字母表示差異顯著(P<0.05);PFG1、PFG2、PFG3和PFG4代表功能群 本實驗按相對重要值比重(占各自功能群重要值80%—90%)取4個功能群主要物種來研究種間關系(表4)。在4個功能群中,重要值最大的分別是麻櫟(3.25)、賽山梅(1.29)、木荷(19.35)、毛柄連蕊茶(7.79)。 表4 午潮山4個功能群主要物種的重要值和生態位寬度 使用Levins(BL)和shannon(BS)指數測量生態位寬度,對環境適應性強的物種具有較大的生態位寬度。午潮山4個功能群主要物種的生態位寬度存在較大差異,Levins和Shannon指數測定結果差異不大,BL和BS的變化范圍分別為9.45—62.32和2.27—4.18。其中檵木的生態位寬度最大,其BL和BS分別為62.32和4.18,隨后是老鼠矢(Symplocosstellaris)(59.80和4.15)和木荷(58.52和4.14),濕地松最小(9.45和2.27)。重要值和生態位寬度成正比,但是也有例外,如PFG1中的濕地松重要值雖然大,但是其生態位寬度卻很小,而白櫟(Quercusfabri)重要值只有1.70但是有較高的生態位寬度(39.32和3.76)。不同功能群的物種數值反差更加明顯。 午潮山森林群落4個功能群主要物種的Pianka生態位重疊指數(Oik)介于0—1之間,0.6≥Oik>0的有106對,占總對數的55.8%,Oik>0.6的有81對,占總數42.6%(圖2)。Oik=0的僅有3對,麻櫟和檫木、麻櫟和賽山梅、濕地松和賽山梅。高生態位重疊值的物種對,通常有相對高的生態位寬度和重要值。PFG1物種與其他物種之間的生態位重疊程度普遍較低。高生態位重疊指數主要集中于除PFG1功能群之外的物種之間,其中,杉木和白花龍與其他樹種生態位重疊指數大多低于0.6。隨著演替進行,同一功能群物種間的生態位重疊值增加。 圖2 午潮山4個功能群主要物種生態位重疊指數矩陣圖Fig.2 Correlation matrix of the Niche overlap of the dominant species in four functional groups in Wuchao Mountain χ2檢驗可以準確的反映種間聯結的顯著度,且提供了量化顯著度的參數。由于木荷出現在所有樣方中以及算法的缺陷,χ2統計量為無限大,因此木荷的χ2檢驗結果不能考慮在內。除去木荷,共171個種對(圖3),80對為正聯結,占46.8%;84對負聯結,占49.1%;7對沒有聯結。正負聯結比為0.95,正相關為顯著和極顯著的分別為4和10,占2.3%和5.8%;負相關為顯著和極顯著的分別為3和14,占1.8%和8.2%。結果與總體聯結性呈負聯結一致,聯結程度較弱。 圖3 午潮山3個功能群主要物種χ2指數矩陣圖Fig.3 Correlation matrix of the χ2 text results of the dominant species in three functional groups in Wuchao Mountain3.841≤χ2<6.635:物種間顯著正聯結(0.01 DI指數被用于定量表達種間聯結性。190對物種中,112對(58.9%)表現為極強聯結,24對(12.6%)表現為強聯結(圖4)。此外,42對為弱聯結,12對無聯結,分別占總物種對的22.1%和6.3%。種間聯結的結果與生態位重疊基本一致,強聯結的種對和高重疊的生態位代表了相似的環境需求。 圖4 午潮山3個功能群主要物種DI指數矩陣圖Fig.4 Correlation matrix of the Dice index (DI) results of the dominant species in three functional groups in Wuchao MountainDI<0.2:無聯結;0.2≤DI<0.4:弱聯結;0.4≤DI<0.6:強聯結;0.6≤DI:極強聯結 Spearman秩相關系數可以用來分析物種協方差(圖5)。56對物種(29.5%)表現為極顯著正相關,10對(5.3%)表現為顯著正相關。此外,55對物種(28.9%)表現為極顯著負相關,9對(4.7%)表現為顯著負相關,60對不相關。此結果補充了木荷與其他物種的聯結性,且正負相關種對的比值為1.03,與χ2檢驗結果基本一致,但是種間顯極著聯結的對數明顯多于χ2檢驗,這是由于χ2檢驗不能準確表達物種間聯結差異性[35]。與生態位重疊結果類似,演替后期物種間的聯結強度要大于演替早期物種。 圖5 午潮山3個功能群主要物種Spearman秩相關系數矩陣圖Fig.5 Correlation matrix of the Spearman′s rank correlation coefficient of the dominant species in three functional groups in Wuchao Mountainrs≥0.1492:極顯著正相關(P≤0.01);0.1492>rs≥0.1133:顯著正相關(0.01 本文應用k-均值聚類法根據物種的葉經濟譜和木材密度等11個功能性狀將其客觀地劃分為4類功能群,并分析不同功能群間的性狀差異定義其演替階段。隨著次生演替的進行,植物從大比葉面積、葉片氮磷鉀含量高的資源獲取性策略向比葉面積小、葉片氮磷鉀含量匱乏的保守型策略轉化;關于葉經濟譜的研究結果同樣證明了這一點[45]。同時,本實驗觀察到采取積極生存策略的演替早期樹種具有低木材密度、喜陽、短壽命的特征,與之前的研究一致[17]。4個功能群有效地體現了對不同生態環境響應和生態系統功能貢獻的植物組合,這為深入研究植物種間關系和群落演替趨勢提供框架基礎[15, 46- 47]。 群落是物種連續更迭的結果,這些物種被環境篩選,同時又構成了新的環境[48]。演替初期,人工種植的杉木和濕地松以及自然更新的麻櫟等先鋒種改變了植被毀壞的午潮山地區的環境條件。隨著演替進行,木荷[49]、青岡、毛柄連蕊茶和薄葉山礬等中性或耐陰樹種逐漸增多,而喜陽的針葉樹種更新緩慢甚至死亡[50]。經過不同種群的長期競爭和適應,最終形成了午潮山以木荷為優勢種的森林群落。 物種對環境資源的利用程度可以通過物種生態位和種間關系的變化格局體現。麻櫟和濕地松等樹種(PFG1)有較高的重要值,主要由于其均為大徑材,對周圍其他樹種幼苗有較大影響。但是,這些喜陽的先鋒樹種擁有較窄的生態位寬度且缺乏幼苗,意味著它們適應和獲取當前生境資源的能力要弱于其他物種,生長更新緩慢。PFG2、PFG3中大多是耐陰適陽的喬木樹種,如小葉石楠、木荷和青岡等,它們生態位寬度較大,說明在群落中分布較廣,對環境資源有較強的利用能力[24]。PFG4中的耐陰灌木擁有大生態位寬度,側面反映了當前生境是闊葉林密閉樹冠下的濕冷生境。生態位重疊程度越低表示物種對相似資源的需求越低,反之,則代表種間競爭激烈[51]。從資源競爭和利用的角度來看,耐陰樹種均擁有大生態位寬度和生態位重疊,表明它們具有較強地適應當前生境的能力以及需求相似生境資源的生存策略。Gu等研究了不同演替動態次生林的生態位和種間聯機,發現隨著演替進行,耐陰樹種與其他樹種的生態位重疊值隨之增加[19]。綜上,隨著不同種群的長期競爭和適應,午潮山森林群落正向后期演替階段進行。 群落結構和物種組成的穩定性會隨著群落演替不斷上升,越接近頂級群落越穩定[52- 53]。本實驗中,主要物種的整體聯結性和正負聯結比均顯示種間水平以負聯結為主或與正聯結持平,但是顯著正聯結種對占優勢。負聯結種對較多可能是由于濕地松作為樣地中的外來種,胸徑均超過20 cm,生長速度快且對資源占有能力較強,與大部分樹種都成顯著負聯結。有研究指出,正負聯結比小于1的情況除了次生演替還有可能是人為連續干擾造成的[54]。午潮山在1960年作為科研基地后,人為砍伐和偷盜樹木的行為被禁止,而1992年后,其作為國家森林公園開放,次生演替明顯受到周邊人為活動的影響,而這些因素是如何影響森林群落結構還需要進一步的調查和研究。 不同功能群物種的種間聯結強度不同,而極顯著的種間聯結物種對達到了半數以上,要高于一些亞熱帶常綠闊葉林的結果[55]。正聯結的物種對意味著分享相似資源同時擁有較寬的生態位重疊,如石櫟(Lithocarpusglaber)和毛柄連蕊茶,這與之前的研究一致[19, 24, 56]。負聯結的種對說明物種有不同的環境需求,例如,青岡在林下形成的環境可供毛柄連蕊茶等耐陰物種生長,但是不適合檫木幼苗這類喜光的物種;早期喜陽先鋒種(PFG1)與其他功能群物種均處于弱強度負聯結,在群落中生態位寬度和生態位重疊均比較低。以上提到的物種間負聯結的原因屬于生境差異的范疇,而類似PFG3和PFG4之間的種間關系則是由于資源競爭導致的負聯結[22, 56],如木荷和毛柄連蕊茶幼苗對林下的資源競爭激烈。因此,在演替中后期,資源競爭是物種競爭的主要原因。 盡管人工種植針葉樹種能加快群落早期演替,但也限制演替中后期(PFG3和PFG4)的樹種更新。人類活動同樣影響森林演替和群落組成結構,如森林片段化和邊緣效應會影響種子的傳播機制[57]。在此,本文針對午潮山森林群落的演替和結構穩定性,結合實驗結果為森林管理提供一些意見。 首先,自然演替需要相當漫長的時間,選擇性的砍伐先鋒種和人為補植演替中后期的物種是加速林分演替不錯的方式。考慮到一些有經濟和景觀價值的先鋒種(如檫木),在林冠層間隙或者與耐陰樹種(如窄基紅褐柃)成正聯結搭配種植,既充分利用了有限的環境資源又維護了生物多樣性和群落穩定性。此外,白櫟、馬銀花和杉木等價值低的樹種,有較大的生態寬度并與青岡、木荷等分享相似的資源,適當伐除可釋放供中后期樹種的生長資源。從而推動中后期目的樹種更新演替,加速午潮山的森林群落向頂級群落發展。其次,濕地松作為外來物種雖然在造林初期生長快、成活率高[58],但是在森林演替中后期與其他物種成負聯結,減緩群落演替進程。根據文本結論,可在濕地松周圍搭配種植白花龍和木荷等喜光的植物,減少濕地松造成的開放地。 最后,本研究在關于生態系統功能和群落結構的基礎上明確了功能群對理解物種利用資源的作用及其對森林經營的實際意義,為分析和指導次生林更新恢復提供了實用的框架,而生態位和種間聯結為生境和資源利用提供詳細的指示。未來的研究還需改善這種方法,如選用更加有代表性的性狀或者能更全面展示物種相互關系的方式,以期有效地提升森林結構和經營模式。1.6 數據分析
2 結果
2.1 功能群的劃分及性狀差異
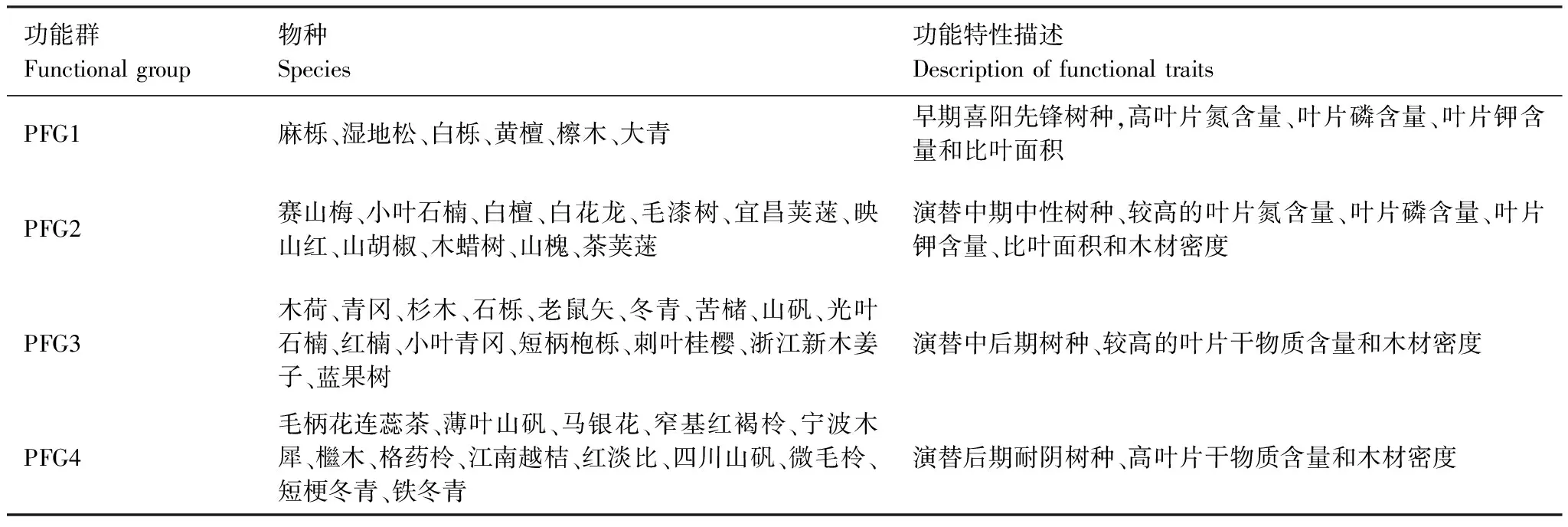
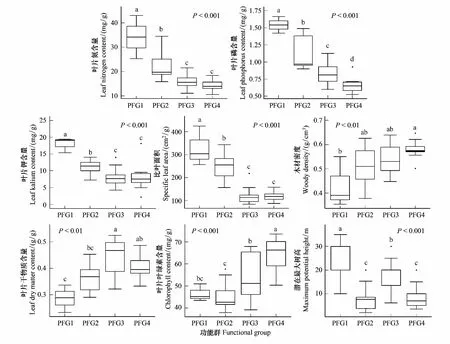
2.2 重要值和生態位寬度
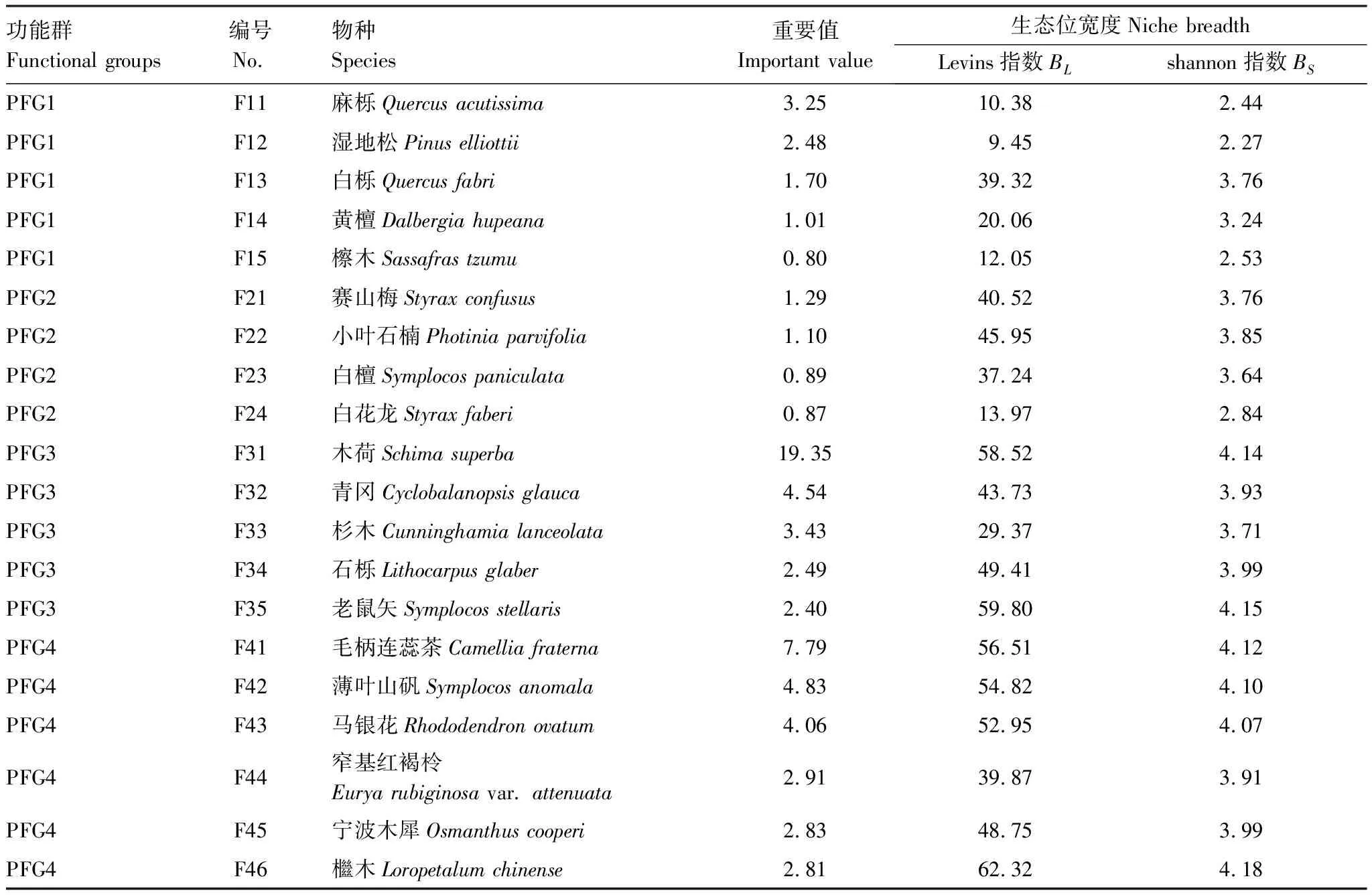
2.3 生態位重疊

2.4 物種的總體聯結性
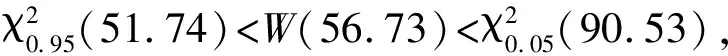
2.5 種對間的聯結性
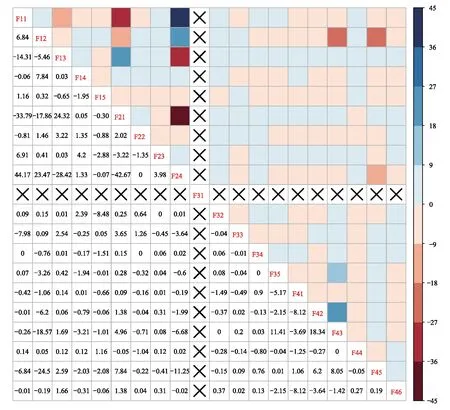
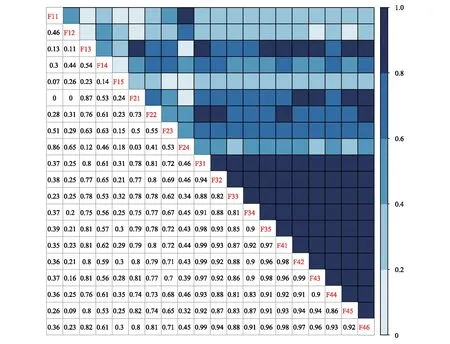
3 討論
3.1 午潮山植物功能群劃分
3.2 午潮山次生林的演替趨勢
3.3 午潮山優勢物種的種間競爭
3.4 森林管理建議
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
領導文萃(2015年4期)2015-02-28 09:19:05
