全外顯子組測序揭示家族性滲出性玻璃體視網膜病變新型LRP5突變的研究
2021-06-04 07:57:42趙汝蓮李姝錦楊正林
實用醫院臨床雜志 2021年3期
關鍵詞:信號
陳 晨,趙汝蓮,彭 麗,李姝錦,4,楊正林,,4
(1.中國科學院成都生物研究所,四川 成都 610041;2.中國科學院大學,北京 100049;3.電子科技大學,四川 成都 611731;4.四川省醫學科學院·四川省人民醫院,四川 成都 610072)
家族性滲出性玻璃體視網膜病變(FEVR)是一種致盲性疾病,是導致青少年視力損傷的主要原因之一,最早由Criswick和Schepens報道[1]。在本研究中,通過對兩個FEVR家族的先證者進行全外顯子測序分析以及Sanger測序驗證,成功鑒定了兩個新的錯義突變:c.3503A>G;p.Y1168C和c.4391T>C;p.M1464T。經過熒光素酶報告基因實驗,發現本研究鑒定出的兩個LRP5基因的新錯義突變均使Norrin/β-catenin信號通路活性顯著下調,這可能導致FEVR的發生。本研究揭示了FEVR患者的遺傳因素,擴寬了LRP5基因的突變譜,為FEVR的分子診斷提供理論依據和新的靶點,從而指導臨床。
1 對象與方法
1.1 研究對象選取2019年4月收集的樣本資料。罹患FEVR的兩個不相關家庭均來自印度Aravind眼科醫院。所有受試者均已簽署書面知情同意,所有實驗均在赫爾辛基宣言的宗旨指導下進行。該研究得到四川省人民醫院倫理監督委員會的批準。通過B超、裂隙燈生物顯微鏡、眼底照相和眼底熒光素血管造影術(FFA)進行FEVR的診斷。所有患者均排除早產,吸氧史或吸毒史。
1.2 DNA提取和全外顯子組測序使用DNA提取試劑盒(TianGen,北京,中國)從外周血樣本中提取先證者及其家系的基因組DNA樣本。家系先證者及相關家系成員的外周血DNA于2019年4月提取完成,存放于-80 ℃,用于后續全外顯子測序。使用全外顯子組測序技術對兩個先證者的DNA樣本測序。DNA文庫的構建基于Agilent SureSelect Human All Exon V5試劑盒(Agilent Technologies,Santa Clara,CA,USA)。文庫測序基于HiSeq2500測序儀(Illumina,圣地亞哥,CA,美國)。原始數據與UCSC hg19(http://genome.ucsc.edu)比對基于BWA(http://bio-bwa.sourceforge.net/)。SAMTOO LS(http://samtools.sourceforge.net/)和ANNOVAR分析用來識別SNP以及插入和刪除(indels)。使用以下數據庫對突變進行次要等位基因頻率(MAF)過濾(MAF <0.001):gnomAD數據庫(http://gnomad.broadinstitute.org/),1000 genomes數據庫(www.internationalgenome.org),dbSNP138數據庫(www.ncbi.nlm.nih.gov/projects/SNP),以及由1600個通過全外顯子組測序(WES)的對照樣本生成的內部數據庫[2~5]。候選致病突變經在線工具(SIFT/PROVEAN和Variant Effect Predictor)預測候選變體是否有害。所有候選致病突變均符合常染色體顯性遺傳模式。
1.3 突變驗證2020年5月,對所有可獲得的家庭成員實施Sanger測序,以驗證候選突變是否與FEVR表型共分離。使用Primer3在線工具(https://bioinfo.ut.ee/primer3-0.4.0/)設計聚合酶鏈反應(PCR)引物。引物的合成由生工生物工程(中國上海)承擔。使用以下引物擴增基因組DNA片段:F:5′-CTGAGAGGCAGGGGCTTT-3′,R:5′-CGCTCACCCTCTCTGAGACT-3′。PCR產物使用FastAP熱敏堿性磷酸酶(Thermo Scientific Fermentas)純化。
1.4 質粒定點誘變和熒光素酶測定(Luciferase assay)將野生型LRP5基因編碼序列克隆到C末端帶有HA標簽的pCDNA3.1載體中。使用QuickChangeR Lightning Site-directed Mutagenesis Kit (New England Biolabs,MA,USA)試劑盒進行DNA定點突變,構建攜帶c.3503A>G;p.Y1168C和c.4391T>C;p.M1464T定點突變。使用EntransterTM-H4000轉染試劑(Engreen,中國北京),將100 ngNorrin,100 ngFZD4、200 ngpGL4.1與100 ng野生型或攜帶突變的LRP5質粒或Vector空載體共轉染進HEK293 STF細胞。48 h后,去除培養基后,利用1×PBS清洗細胞,加入150 ml細胞裂解液于冰上裂解5 min,待細胞裂解完全后將裂解產物移至1.5 ml離心管,12000×g離心后吸取100 ml上清液于96孔板上。通過雙重熒光素酶報告基因測定系統(TransGenBiotec)測量報告基因活性,相對熒光素酶單位(螢火蟲熒光/海腎熒光活性)即代表Norrin/β-catenin信號通路的活性強度。
1.5 細胞培養和免疫熒光染色(ICC)將COS7細胞接種在24孔板(Corning)中的細胞爬片上,于5%二氧化碳培養箱中37 ℃培養。轉染野生型LRP5質粒、或突變型LRP5質粒、或Vector空載體。48 h后,在室溫下于4%PFA(多聚甲醛)中固定20 min。隨后,將爬片在磷酸鹽緩沖溶液(PBS)(Sigma)中潤洗3次(5分鐘/次)。在室溫下用封閉緩沖液(5%FBS、0.2%Triton X-100配制于PBS中)封閉30分鐘,隨后加入一抗,于4 ℃搖床上孵育過夜。次日于室溫下,用熒光標記的二抗孵育1小時。通過激光共聚焦顯微鏡800(Zeiss,德國)拍攝免疫熒光圖像。
1.6 抗體免疫熒光染色使用的抗體為:anti-HA(Roche,1∶2000稀釋),DAPI(Cell Signaling Technology,1:500稀釋),Alexa FluorTM-488 goat Anti-Rat(Invitrogen,1∶500稀釋)。
1.7 統計學方法使用GraphPadPrism 6.0軟件對熒光素酶報告基因實驗結果進行統計分析。計數資料比較采用Tukey檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 臨床特征本研究招募了兩個罹患FEVR的印度家系。第一個家系,先證者于50歲時確診FEVR。第二個家系,先證者于18歲時確診患有FEVR,其父親和妹妹均被診斷為FEVR。
2.2 LRP5基因的新突變使用全外顯子測序(WES),和生物信息學分析,在兩個印度FEVR家系中初步篩選出了兩個新的LRP5突變:c.3503A>G;p.Y1168C和c.4391T>C;p.M1464T。在dpSNP138、Exome Variant Server、ExAC和gnomAD中未觀察到這兩個突變。此外,與內部數據庫(1600名健康對照的外顯子組測序分析數據)對比并未觀察到上述兩個突變。由此推斷c.3503A>G;p.Y1168C和c.4391T>C;p.M1464T是FEVR致病候選突變。
2.3 Sanger測序驗證結果為了驗證全外顯子測序結果的準確性,我們利用Sanger測序和共分離分析,進一步確定突變的致病可能性。對于c.3503A>G;p.Y1168C突變,先證者被鑒定為純合子攜帶者,其三個兒子均為雜合子攜帶者(圖1a)。此外,先證者的丈夫不攜帶LRP5突變(圖1a)。譜系分析表明c.3503A>G;p.Y1168C突變與該家族的疾病表型共分離,得出結論,該突變以常染色體顯性方式遺傳。對于c.4391T>C;p.M1464T突變,先證者、其父親及妹妹均被診斷出患有FEVR并攜帶雜合突變(圖1b),然而,本研究缺乏先證者母親的遺傳數據。家系分析表明c.4391T>C;p.M1464T突變與該家族中的疾病表型共分離,得出結論,該突變以常染色體顯性方式遺傳。
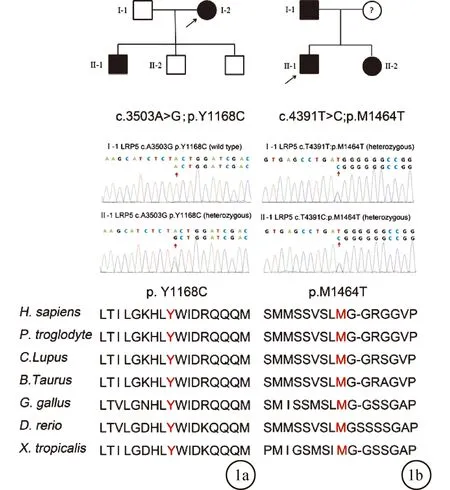
圖1 家系遺傳圖譜、Sanger測序結果及物種保守性分析結果 a:家系1;b:家系2
2.4 LRP5基因的突變使Norrin/β-catenin信號通路下調LRP5是激活Norrin/β-catenin信號傳導通路的重要組分。為了研究突變對Norrin/β-catenin信號傳導的影響,在HEK293STF細胞中進行了熒光素酶報告基因檢測。將NDP、FZD4、PGL4.1(組成型啟動子下的海腎螢光素酶作為內部對照)與野生型LRP5、突變型LRP5或Vector空載體共轉染,在細胞裂解液中依次加入螢火蟲熒光素酶底物和海腎熒光素酶底物,經檢測后,兩次測得的結果比值即為Norrin/β-catenin信號通路的相對活性。與野生型質粒相比,兩個突變均顯著降低LRP5激活Norrin/β-catenin信號通路活性的能力,差異有統計學意義(P<0.05),見圖2a。
2.5 突變并不影響LRP5蛋白的定位使用細胞免疫熒光化學(ICC)研究突變是否引起LRP5蛋白的定位改變。結果表明這兩個突變蛋白與野生型LRP5蛋白定位一致,表達于細胞質中,且主要積累于高爾基體中,見圖2b。
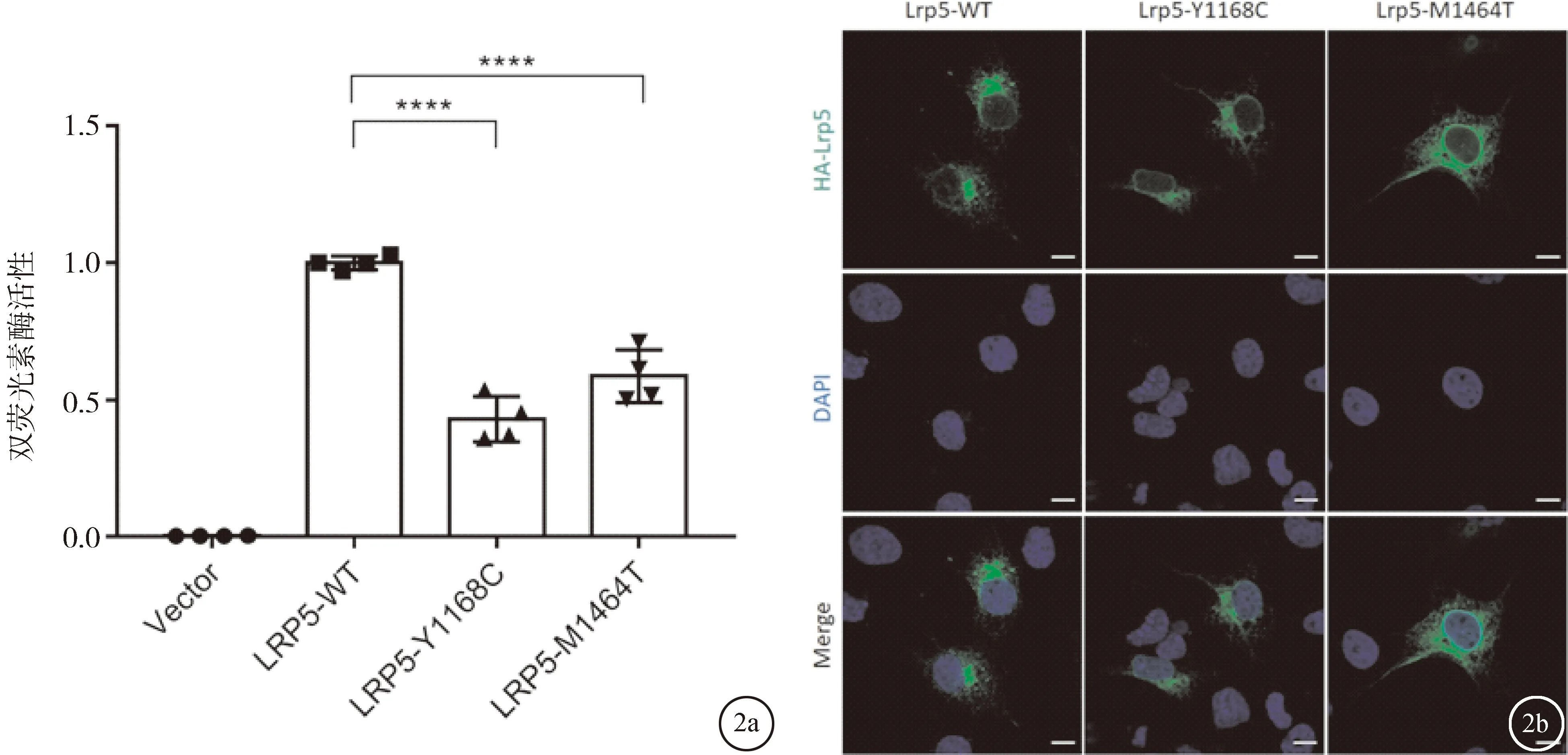
圖2 雙熒光素酶報告基因實驗結果及ICC結果 a:實驗結果,**** P<0.0001;b:ICC結果,比例尺:25 μm
3 討論
家族性滲出性玻璃體視網膜病變(FEVR)是一種罕見的遺傳性視網膜血管疾病,是青少年視力喪失的主要原因。目前,由于FEVR的臨床和遺傳異質性,其診斷仍存在困難。FEVR具有高度的臨床異質性,其臨床表現多樣。典型的FEVR可表現為周圍血管缺失、視網膜新血管生成、視網膜牽拉和脫離、玻璃體出血和黃斑性近視,最終導致失明[6]。FEVR具有遺傳異質性即其遺傳方式多樣,包括常染色體顯性遺傳、常染色體隱性遺傳或X性染色體連鎖遺傳[6,7]。到目前為止,NDP、FZD4、LRP5、TSPAN12、ZNF408、KIF11、RCBTB1、CTNNB1、ILK、JAG1、CTNNA1、ATOH7和11p12-13染色體上的EVR3[8~16]的突變都可能導致FEVR的發生。盡管在最近幾十年中,研究者們已陸續鑒定出FEVR致病相關的新基因及其突變,但迄今為止報道的突變僅能解釋約50%的FEVR病例。其中,由NDP、FZD4、LRP5、TSPAN12和CTNNB1基因編碼的蛋白質參與Norrin/β-catenin信號傳導途徑,在視網膜血管發育中起著至關重要的作用[17]。Norrin/β-catenin信號傳導途徑是經典Wnt信號通路,在生物的生長過程中起重要作用。Norrin/β-catenin信號通路的正常活性對視網膜血管形成過程尤為重要。Norrin/β-catenin信號通路的幾個核心組成部分的突變均會導致到FEVR的發生。
在本研究中,通過全外顯子測序技術在兩個印度家庭中的先證者NDA樣本中鑒定了兩個與FEVR相關的LRP5基因的新突變:c.3503A>G;p.Y1168C和c.4391T>C。利用Sanger測序在家系的成員中驗證得到的候選致病新突變。臨床診斷結果以及Sanger測序結果表明這兩個新突變的基因表型共分離,并且突變所累及的氨基酸殘基在各物種中具有高度保守性。結論:這兩個突變均以常染色體顯性方式遺傳,見圖1。
接下來對c.3503A>G;p.Y1168C和c.4391T>C突變的致病性進行了研究。雙熒光素酶活性檢測可以反映Norrin/β-catenin信號通路的活性。實驗發現,與野生型LRP5蛋白相比這兩個新突變均導致Norrin/β-catenin信號通路受到抑制,見圖2a。這說明c.3503A>G;p.Y1168C和c.4391T>C突變是致病突變。接著探討了突變對蛋白定位是否有影響。細胞免疫熒光實驗是常用的觀察突變對蛋白定位影響的實驗手段。如圖2b所示,與野生型LRP5相比,突變并沒有影響蛋白的定位。野生型及突變蛋白均表達于細胞質中。這表明LRP5蛋白的定位并未受到這兩個突變的影響。
綜上,本實驗發現了c.3503A>G;p.Y1168C和c.4391T>C;p.M1464T兩個FEVR致病突變。本研究擴大了LRP5基因突變譜,為FEVR的遺傳診斷和咨詢提供了理論依據。然而,突變導致FEVR的致病機制仍有待進一步研究。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
