懷玉山高山馬鈴薯蔗糖合酶基因的克隆和分析
2021-05-31 00:40:56洪森榮鄧雨晴吳洪婷郭淑貞楊于萱陳榮華
福建農業學報 2021年2期
洪森榮,鄧雨晴,吳洪婷,陳 婷,郭淑貞,楊于萱,蔡 紅,陳榮華
(1. 上饒師范學院生命科學學院,江西 上饒 334001;2. 上饒市藥食同源植物資源保護與利用重點實驗室,江西 上饒 334001;3. 上饒市薯芋類作物種質保存與利用重點實驗室,江西 上饒 334001;4. 上饒農業技術創新研究院,江西 上饒 334001;5. 上饒市紅日農業開發有限公司,江西 上饒 334700)
0 引言
【研究意義】馬鈴薯(Solanum tuberosum L.)為茄科茄屬(Solanum L.)雙子葉一年生植物,原產于南美洲安第斯山區,是全球僅次于水稻、小麥、玉米的第四主糧作物,是全球最重要的塊莖類作物,也是重要的糧菜兼用作物和工業原料[1?2]。馬鈴薯營養豐富,可實現周年供應,可作糧用、菜用、飼用[3]。玉米、小麥、水稻易受限于氣候、水肥等條件,而馬鈴薯在偏遠缺水地區仍可實現豐產,且產業鏈長,開發前景廣闊,是我國脫貧攻堅的助力產業[3]。中國是第一大馬鈴薯生產國,種植面積和產量均居世界首位,在緩解我國食物壓力、保障我國糧食安全方面作用突出[3]。高山馬鈴薯食用價值和營養價值高,管理成本和種植技術低,且高山馬鈴薯對高山地區環境適應能力強,可在高海拔山區推廣種植,具有較強的產業化潛力[4]。江西懷玉山屬于高海拔高山地區,紫外線強烈,晝夜溫差大,馬鈴薯品種應選擇早熟型,如懷玉山高山馬鈴薯(S. tuberosum L.cv. Huaiyushan)[5]。懷玉山高山馬鈴薯,又稱麻籽洋芋,國家地理標志農產品(2013年)。懷玉山高山馬鈴薯藥食兼優。食用,形橢圓,具麻點,色黃肉白(屬于外觀品質),營養豐富,切后不黑,味甘爽口,實為一道天然美味佳肴;藥用,不含還原糖,富含膳食纖維,可和中養胃、健脾利濕、降糖降脂、美容養顏、防治胃癌[6]。蔗糖是運輸和貯存光合碳水化合物的主要形式,是植物糖代謝的中心調控樞紐[7]。蔗糖合酶(Sucrose synthase,SuSy)可分解蔗糖,也可合成蔗糖,具有雙重屬性[8],主要參與植物韌皮部代謝、碳源分配、纖維素和淀粉合成,還可影響植物抗逆性、種子發育、果實品質形成、生殖生長、糖信號轉導以及生物固氮能力,為植物體生長發育提供碳源和能量動力[9]。因此,分析懷玉山高山馬鈴薯蔗糖合酶的核酸序列信息,預測其蛋白結構與功能,對進一步揭示該酶生物學功能具有重要指導意義。【前人研究進展】目前已在蘋果[10]、苦蕎[11]、毛白楊[12]、木薯[13]和白及[14]等多種高等植物中分離得到了蔗糖合酶基因。而關于懷玉山高山馬鈴薯的研究主要集中于全基因組重測序分析[15]、轉錄組分析[5]、DNA甲基化MSAP分析[6]、遺傳多樣性同工酶分析[16]、脫毒方法[17]和病毒DAS-ELISA檢測與分析[18]等研究。【本研究切入點】但有關懷玉山馬鈴薯蔗糖合酶基因結構與功能的研究還未見報道。【擬解決的關鍵問題】本研究通過從構建的懷玉山馬鈴薯轉錄組數據庫中挑選蔗糖合酶基因序列(SuSy 4,PDSC Gene ID:PGSC0003DMG400002895,PDSC Transcript ID:PGSC0003DMT400007506,iTAG Transcript ID:Sotub12g008670.1.1,NCBI Reference Sequence/GenBank:M18745.1,Locus At:At3g43190),利用在線生物信息學分析軟件分析蔗糖合酶基因序列的氨基酸序列、理化性質、蛋白二級結構、結構功能域、三級結構、亞細胞定位,為分析蔗糖合酶保守結構域提供基礎資料,為馬鈴薯蔗糖合酶的進一步研究提供生物信息學方面的參考。
1 材料與方法
1.1 材料和試劑
懷玉山高山馬鈴薯試管苗(上饒師范學院生命科學學院植物組織培養室提供)、Trizol總RNA提取試劑、PCR反應2×GoldStarTaqMasterMix購自北京康為世紀公司;M-MLV cDNA第一鏈合成試劑、PrimeScript?RT Reagent Kit With gDNA Eraser(Perfect Real Time)、PMD18-T vector及SYBR? Premix Ex TaqTM II均購自TaKaRa公司;Gel Extraction Kit購自Omega公司;引物由上海生工生物工程有限公司合成。大腸桿菌(E. coli)DH5α 由本實驗室保存;其 他常規試劑均為國產分析純試劑。
1.2 試驗方法
1.2.1 總RNA的提取和cDNA第一鏈的合成 用Trizol試劑提取懷玉山高山馬鈴薯試管苗的總RNA,提取步驟按說明書進行,使用紫外分光光度計和瓊脂糖凝膠電泳檢測RNA的濃度和完整性。以提取獲得的RNA為模版,按照M-MLV cDNA第一鏈合成試劑盒說明書合成cDNA第一鏈。逆轉錄引物用Oligo(dT)18 Primer:5′-GGCCACGCGTCGACTA GTACTTTTTTTTTTTTTTTTTT-3′,具體步驟按說明 書進行。
1.2.2 懷玉山高山馬鈴薯蔗糖合酶基因的克隆 利用懷玉山高山馬鈴薯試管苗轉錄組數據庫篩選到蔗糖合酶基因的核心片段(CDS全長),運用Primer Premier 5.0設計基因特異性引物(F:ATGGCTGAA CGTGTTCTGACTC;R:TCACTCAGCAGCCAATGGA AC)。PCR擴增條件:95 ℃ 2 min;95 ℃ 30 s,55 ℃30 s,72 ℃ 2.5 min,35個循環;72 ℃ 10 min。PCR產物經1%瓊脂糖凝膠電泳檢測后,將含有目的基因的條帶與pMD19-T載體連接并用熱激法轉化到感受態細胞E.coli DH5α,經鑒定正確的陽性轉化子提取質 粒送往上海生工進行測序。
1.2.3 懷玉山高山馬鈴薯蔗糖合酶基因的生物信息學分析 按照張林等[19]的方法對懷玉山高山馬鈴薯蔗糖合酶進行氨基酸序列分析、理化性質分析、結構預測、功能分析。使用BioEdit軟件翻譯4個基因序列為氨基酸序列,用ProtParam(https://web.expasy.org/protparam/)預測蛋白的理化性質,用ProtScale(https://web.expasy.org/cgi-bin/protscale/protscale.pl)預測蛋白疏/親水性。使用GOR IV軟件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_gor4.html)在線預測蛋白質二級結構。使用SWISS-MOLD(http://www.swissmodel.expasy.org/interac)在線預測蛋白的三級結 構。
2 結果與分析
2.1 懷玉山高山馬鈴薯RNA提取
懷玉山高山馬鈴薯試管苗總RNA的電泳圖見圖1。RNA條帶清晰,無色素、蛋白、糖類等雜質污染,28/23S亮度大于18/16S,濃度為602.00 ng·μL?1,總量為18.06 μg,RIN值=8.4,OD260/280=2.17,OD260/230=2.22,表明提取的RNA較為完整,質量較高,可以進 行后續實驗。
2.2 懷玉山高山馬鈴薯蔗糖合酶基因cDNA序列
通過PCR擴增技術(圖2),懷玉山高山馬鈴薯蔗糖合酶基因cDNA總長度為2 418 bp(圖3),G+C 含量為45.08%。
2.3 懷玉山高山馬鈴薯蔗糖合酶蛋白氨基酸序列

圖 1 懷玉山高山馬鈴薯總RNA的電泳Fig. 1 Electrophoresis of total RNA from Alpine potato of Huaiyushan

圖 2 懷玉山高山馬鈴薯蔗糖合酶基因PCR擴增Fig. 2 PCR amplification of SuSy gene in Alpine potato of Huaiyushan
Protparam預測顯示懷玉山高山馬鈴薯蔗糖合酶蛋白氨基酸序列見圖4。懷玉山高山馬鈴薯蔗糖合酶蛋白由805個氨基酸組成,分子量92 471.33 Da,等電點5.87,為親水性蛋白。Leu、Glu、Val、Ala、Lys、Ile、Gly和Arg是懷玉山高山馬鈴薯蔗糖合酶SuSy氨基酸序列中含量最為豐富的氨基酸,其平均含量分別為11.1%、9.3%、6.6%、7.0%、6.3%、6.0%、5.7%和5.5%,不含Pyl和Sec(表1)。帶負電殘基總數(Asp+Glu)為114,正電荷殘基總數(Arg+Lys)為95。估計半衰期為30 h(哺乳動物網織紅細胞,體外),>20 h(酵母,體內),>10 h(大腸桿菌,體內)。哺乳動物與酵母、大腸桿菌酶的半衰期差異較大,主要受氨基末端氨基酸殘基(如Met、Ala、Glu等)影響。懷玉山高山馬鈴薯蔗糖合酶蛋白失穩指數(II)計算為33.96,組氨酸(His)、天冬氨酸(Asp)、精氨酸(Arg)、谷氨酸(Glu)、賴氨酸(Lys)5種氨基酸殘基構成酶的活性中心,與酶的穩定性相關,表明懷玉山高山馬鈴薯蔗糖合酶蛋白分類為穩定的。

圖 3 懷玉山高山馬鈴薯蔗糖合酶基因堿基組成Fig. 3 Base composition of SuSy gene in Alpine potato of Huaiyushan

圖 4 懷玉山高山馬鈴薯蔗糖合酶蛋白氨基酸序列Fig. 4 Amino acid sequence of SuSy in Alpine potato of Huaiyushan
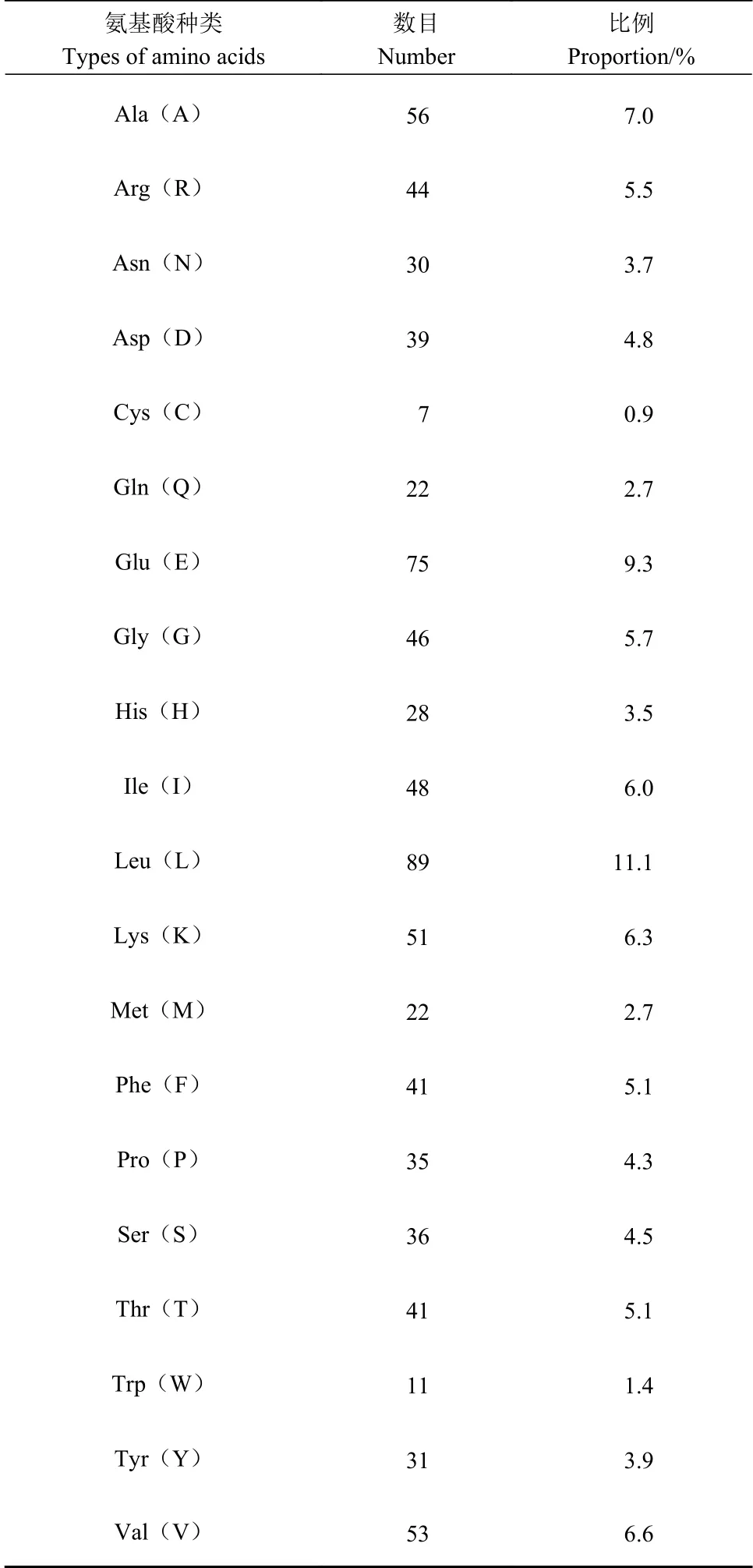
表 1 懷玉山高山馬鈴薯蔗糖合酶蛋白各氨基酸的數目和比例Table 1 Number and proportion of amino acids of SuSy in Alpine potato of Huaiyushan
2.4 懷玉山高山馬鈴薯蔗糖合酶蛋白親疏水性分析
從圖5可知,高峰值(正值)的區域表示疏水的區域,而低谷(負值)的區域是親水區域。疏水性結果分析表明,最大疏水值為2.5左右,在該多肽中說明該處的疏水性最強;親水峰最大值為?3左右,整個蛋白質表現出高度的親水性,說明該蛋白為親水性蛋白質。

圖 5 懷玉山高山馬鈴薯蔗糖合酶蛋白親疏水值分布Fig. 5 Distribution of relative and sparse water values of SuSy in Alpine potato of Huaiyushan
2.5 懷玉山高山馬鈴薯蔗糖合酶二級結構分析
懷玉山高山馬鈴薯蔗糖合酶的二級結構(圖6)預測如下:GOR預測顯示其二級結構由α-螺旋(Alpha helix,Hh,45.84%)、延 伸 鏈(Extended strand,Ee,15.16%)、無規則卷曲(Random coil,Cc,39.01%)構成(圖7)。從分布位點上來看,C端和N端含延伸鏈和α-螺旋,而無規則卷曲、延伸鏈和α -螺旋則散布于整個蛋白質中。
2.6 懷玉山高山馬鈴薯蔗糖合酶三級結構分析
SWISS-MODEL預測顯示懷玉山高山馬鈴薯蔗糖合 酶的三級結構為四聚體(圖8)。
2.7 懷玉山高山馬鈴薯蔗糖合酶亞細胞定位
采用Psort在線軟件對懷玉山高山馬鈴薯蔗糖合酶的表達部位進行預測,結果表明:定位于細胞質中的數量為5,線粒體中的數量為5,葉綠體中的數量為4,表明懷玉山高山馬鈴薯蔗糖合酶主要存在細胞 質、線粒體和葉綠體中。
2.8 懷玉山高山馬鈴薯蔗糖合酶系統進化分析
從構建的進化樹(圖9)中可見,懷玉山高山馬鈴薯與Solanum lycopersicum(番茄)、Solanum pennellii(潘那利番茄)、Solanum chilense(智利番茄)、Solanum tuberosum(馬 鈴 薯)、Capsicum annuum(辣椒)、Capsicum baccatum(風鈴辣椒)等6種植物在一個大分支下,這說明懷玉山高山馬鈴薯蔗糖合酶基因在進化上與六者的親緣關系較近,尤其是與馬鈴薯(Solanum ltuberosum)的進化上具有最高的親緣關系。

圖 6 懷玉山高山馬鈴薯蔗糖合酶二級結構Fig. 6 Secondary structure of SuSy in Alpine potato of Huaiyushan

圖 7 懷玉山高山馬鈴薯蔗糖合酶二級結構各部分的比例Fig. 7 Proportion of various parts in secondary structure of SuSy in Alpine potato of Huaiyushan

圖 8 懷玉山高山馬鈴薯蔗糖合酶三級結構Fig. 8 Tertiary structure of SuSy in Alpine potato in Huaiyushan
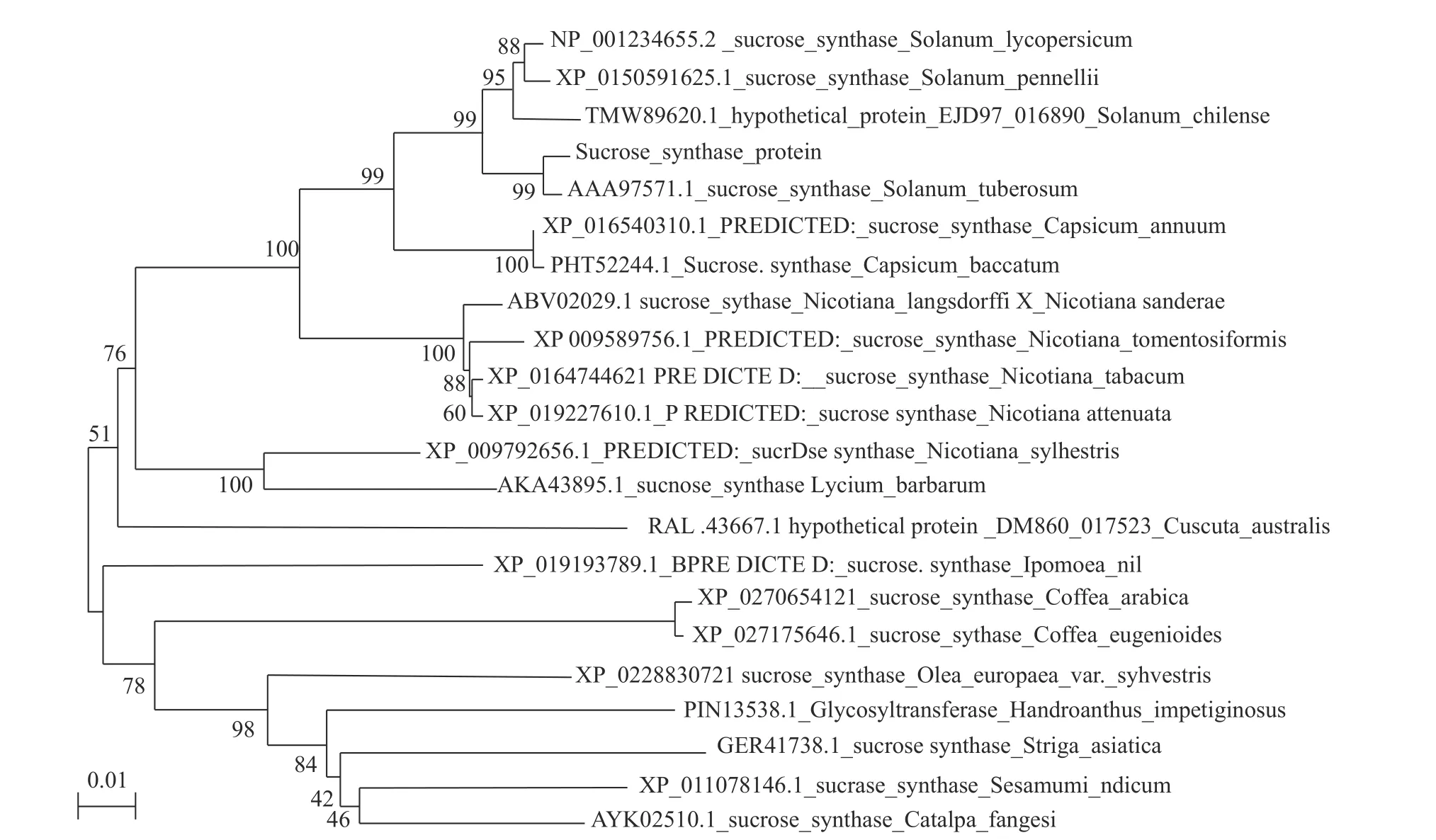
圖 9 懷玉山高山馬鈴薯蔗糖合酶系統進化分析Fig. 9 Phylogenetic analysis of SuSy in Alpine potato of Huaiyushan
3 討論
蔗糖合酶屬于蔗糖代謝的關鍵酶,廣泛存在于植物體內,負責催化蔗糖和UDP-果糖合成UDPG,也負責催化UDPG分解為蔗糖和UDP-果糖的反應[20]。蔗糖合酶與蔗糖代謝其他酶(轉化酶、蔗糖磷酸合酶等)的不同處在于其可催化可逆反應,不需耗能且不需其他酶參與,直接參與植物體內的生物合成及利用[21]。
植物由于長期適應環境得到不斷進化,但執行一定生理功能的基因會顯得相對保守,因此,利用氨基酸序列構建的系統進化分析可確定物種之間的親緣關系,還以此來評估蛋白功能的相關性[22]。本研究對懷玉山高山馬鈴薯(S. tuberosum L. cv.Huaiyushan)蔗糖合酶進行系統進化分析,表明懷玉山高山馬鈴薯(S. tuberosum L. cv. Huaiyushan)與番茄(Solanum lycopersicum)、潘那利番茄(Solanum pennellii)、智利番茄(Solanum chilense)、馬鈴薯(Solanum ltuberosum)、辣椒(Capsicum annuum)、風鈴辣椒(Capsicum baccatum)等6種植物在一個大分支下,這說明該基因在進化上與這六者的親緣關系較近,尤其是與馬鈴薯(Solanum ltuberosum)的進化上具有最高的親緣關系。
蔗糖合酶(SuSy)是蔗糖分解的關鍵酶,被認為是衡量庫強的生化指標,特別是在作物品種中。通過在棉花中表達馬鈴薯Sus基因,發現在Sus高表達系的幼葉中,果糖含量顯著增加,而在伸長纖維中,果糖和葡萄糖含量均增加[23]。在轉基因馬鈴薯中,蔗糖合成酶(Susy)的表達通過引入一個包含Susy編碼區的嵌合基因而改變,Susy活性和淀粉含量增加[24]。隨著對馬鈴薯蔗糖合酶的深入研究,馬鈴薯蔗糖合酶以不同亞型形式存在,這些亞型至少由兩個基因編碼,甚至更多,至今已在45種植物中發現超過80個Sus基因[25]。馬鈴薯蔗糖合成酶基因Sus4-16的5′側翼序列、先導內含子和3′序列的3.6 kb和0.7 kb足以在發育中的塊莖、腋芽和芽的基部組織、分生組織和根冠中誘導高水平表達,并賦予葉片蔗糖誘導能力[26]。在馬鈴薯中發現了兩類不同的蔗糖合成酶基因Sus3和Sus4。根據序列同源性,它們似乎是在雙子葉植物主要科分化之后、番茄和馬鈴薯分化之前進化而來的[21]。這個觀點與本試驗結果一致。也有研究表明,馬鈴薯蔗糖合酶與玉米蔗糖合酶cDNA序列相比,核苷酸序列的同源性約為70%,推導的氨基酸序列約為75%,3個氨基酸區的同源性約為90%[27]。蔗糖合酶通常以二聚體和四聚體的形式存在[28],但四聚體形式存在的蔗糖合酶是有活性的存在形式[29]。研究表明,馬鈴薯蔗糖合酶的N端的細胞靶向結構域(cellular targeting domain,CTD)結構域可以使酶與細胞內的肌動蛋白纖維、線粒體及液泡膜等相結合,參與細胞靶向的調控[30]。利用原位雜交技術研究馬鈴薯淀粉代謝相關基因的表達的影響,結果發現蔗糖合酶基因在塊莖中表達,但在塊莖形成的頂端分生組織或匍匐莖中不表達,編碼顆粒結合淀粉合成酶和淀粉分支酶的mRNAs主要在淀粉質體周圍積累,而編碼蔗糖合成酶的mRNAs沒有顯示出優先分布[31]。本研究采用Psort在線軟件對懷玉山高山馬鈴薯(S. tuberosum L. cv.Huaiyushan)蔗糖合酶的表達部位進行預測,結果表明懷玉山高山馬鈴薯(S. tuberosum L. cv. Huaiyushan)蔗糖合酶主要存在細胞質、線粒體和葉綠體中,并沒有優先分布在某個部位。
4 結論
懷玉山高山馬鈴薯蔗糖合酶具有典型蔗糖合酶的結構特征,其氨基酸序列和核酸序列與同源物種相似度高,在進化上高度保守。
