PP2A調節mbk-2影響秀麗隱桿線蟲胚胎發育
2021-04-12 02:41:38翟佳佳康平
河北醫藥 2021年4期
翟佳佳 康平
秀麗隱桿線蟲(Caenorhabditis elegans,C.elegans)結構簡單、生長周期短、基因與人類基因具有高度同源性、易培養,是研究遺傳、基因組和分子網絡的重要模式。蛋白磷酸酶2A(protein phosphatase 2A,PP2A)屬于絲/蘇氨酸磷酸酶,在調節細胞關鍵功能,如細胞周期、發育、代謝和凋亡中發揮重要作用。mbk-2是秀麗隱桿線蟲雙特異性Yak-1相關激酶家族中的兩種絲氨酸/蘇氨酸激酶之一。mbk-2在早期胚胎發育,特別是在單細胞胚胎的多個微管依賴過程中起重要作用,包括原核遷移、紡錘體定位、微管穩定性和染色體分離[1]。大規模線蟲RNAi篩選發現,增強子pptr-1、pptr-2對mbk-2有調節作用。pptr-1和pptr-2可編碼PP2A的調節亞單位,對調節PP2A功能發揮重要作用。本研究采用秀麗隱桿線蟲mbk-2突變體觀察PP2A調節亞單位pptr-1和pptr-2對mbk-2的調控作用,為進一步研究mbk-2在胚胎發育中的作用及調控機制提供理論依據。
1 材料與方法
1.1 線蟲株系與菌株 野生型線蟲N2、mbk-2基因敲除型線蟲株系、大腸埃希菌OP50 購自美國秀麗線蟲遺傳中心(Caenorhabditis Genetics Center)。
1.2 主要儀器及試劑 氨芐青霉素(上海BBI,A610028);羧芐青霉素(美國Sigma,A6140);L4440質粒(美國Addgene)。體式解剖顯微鏡(廈門Motic);低溫高速離心機(德國eppendorf);激光共聚焦成像系統(德國Leica)。
1.3 實驗方法
1.3.1 線蟲生長培養:線蟲培養采用NGM培養基,稱取NaCl 3 g,瓊脂20 g,蛋白胨2.5 g,溶于975 ml雙蒸水,高壓滅菌。冷卻后,加入1 mol/L CaCl21 ml,5 mg/ml膽固醇1 ml,1 mol/L MgSO41 ml和1 mol/L KPO4緩沖液25 ml,混勻后導入細菌培養皿中。室溫放置2 d后,鋪上OP 50菌,室溫放置2 d后即可培養線蟲。
1.3.2 線蟲同步化:將線蟲常規培養在NGM培養基上,20℃生化培養箱中生長繁殖。用Bleach裂解液沖洗NGM培養基,震蕩,低速離心,吸取上清液,再加入S-basal溶液,震蕩,離心,吸取上清。重復2次加入后加入S-buffer溶液放進入25℃生化恒溫箱中培養過夜。第2天離心,吸取上清,將剩余部分混勻后,涂抹在NGM培養基上.待NGM培養基中的線蟲進入產卵期,將正處于產卵期的線蟲(母蟲)挑到NGM培養基上進行產卵。2~3 h后將線蟲(母蟲)挑走殺死后,將NGM培養基放進25℃的生化培養箱中進行培養。第二天觀察存活情況,發育至成蟲(L4期)后可用來進行相關實驗。
1.3.3 RNAi干擾試驗:RNAi細菌在含有50 g/L氨芐青霉素的LB溶液中于37℃振蕩過夜。將細菌溶液點在NGM瓊脂平板上(含有100 g/L氨芐青霉素、12.5 g/L四環素和1 mmol/L IPTG),并在室溫下誘導細菌過夜。在20℃條件下,將同步化好的N2及mbk-2基因缺陷線蟲在NGM培養基中培養至L4期,喂食RNAi細菌,分別含有沉默pprt-1和pprt-2基因的質粒(沉默組)和L4440空載質粒(對照組)。
1.3.4 孵化率測定:將大約50只線蟲放在裝有300 μl 細菌的RNAi平板上,并對它們進行培養,直到它們長到L4期。然后將線蟲轉移到裝有30 μl RNAi細菌的新鮮瓊脂培養平板(每塊平板上有三只蠕蟲),并將其孵育24 h。然后這些線蟲(新的成蟲)被轉移到另一個RNAi平板上,24 h后移除。將平板上的子代培養24 h后,我們對幼蟲和未孵化的蟲卵數量進行計數。
1.3.5 幼蟲發育分析:線蟲從L1階段喂食RNAi細菌,直到線蟲成年產卵前兩天。預先準備約0.4 mm厚的瓊脂墊,并在使用前與M9溶液和蓋玻片一起放入溫控培養箱中孵育2 h。在蓋玻片上滴加的2 μl M9緩沖液,并在其中解剖8~10個只成蟲,用熔化的凡士林封片,顯微鏡下觀察并拍照記錄。
2 結果
2.1 pprt基因沉默對N2線蟲孵化的影響 采用含pprt-1和pprt-2的RNAi細菌喂養線蟲,觀察抑制pprt對線蟲孵化率的影響,對照組采用含L4440質粒的細菌喂養。結果顯示,抑制pptr-1后,線蟲無蟲卵死亡,所有蟲卵均孵化出幼蟲。而抑制pprt-2后,(39.7±1.53)的蟲卵死亡,其未孵化率為(5.83±0.23%),顯著高于對照組N2和pprt-1干預組。見表1,圖1。
2.2 pprt-1和pprt-2基因沉默對mbk-2突變線蟲孵化的影響 采用pprt-1和pprt-2的RNAi細菌喂養mbk-2突變型線蟲。結果顯示,L4440質粒喂養的N2對照組線蟲,無蟲卵死亡,與N2組相比,mbk-2突變會增加蟲卵的未孵化率,在(1.63±0.51%)。采用RNAi抑制pprt-1后,蟲卵死亡增加,未孵化率(3.73±0.12%),顯著高于N2對照組和pprt-1 RNAi處理組。抑制pprt-2后,線蟲蟲卵的未孵化率達到(76.74±1.62%),顯著高于N2對照組,mbk-2組,mbk-2+pprt-1處理組以及pprt-1RNAi處理組。見表1,圖1。
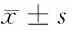
表1 線蟲蟲卵孵化率統計表

圖1 線蟲蟲卵孵化檢測
2.3 pprt-2基因沉默對mbk-2突變線蟲胚胎發育的影響 為研究pprt-2對mbk-2的調控作用,采用pprt-2 RNAi細菌喂養mbk-2突變線蟲。對胚胎發育表型的研究顯示,在單細胞階段,pprt-2 RNAi干預對胚胎發育不產生影響,原核遷移、紡錘體的建立和假裂溝的位置正常。而在雙細胞階段,則表現為明顯缺陷。AB和P1細胞的大小相差很大;當與野生型相比時,AB細胞比P1細胞大得多,和/或在AB細胞和P1細胞中都可以看到多個細胞核。我們還觀察到沒有細胞核或細胞體的額外細胞的存在,這些細胞在許多情況下與其他細胞分離,然后融合回來。AB和P1紡錘體可能已經消失,或者其中一個丟失,或者它們可能有定向缺陷。在一些胚胎中,P1細胞首先分裂,或者AB和P1同時分裂(四極分裂)。其他胚胎表現出極端的胞質分裂缺陷,其中細胞之間或異常的細胞-細胞接觸之間沒有明顯的邊界。或者,當4個單元可以區分時,它們的大小可能有很大的不同,4個細胞有多個細胞核。大多數觀察到的缺陷類似于mbk-2功能的強烈喪失,表明pptr-2影響胚胎此階段的發育需要mbk-2參與。這種相互作用獨有的一種表型是P1細胞分裂的延遲。在ABa和ABp細胞的細胞核形成之后,就在這些細胞分裂之前,P1紡錘體仍然沒有建立。在相同條件下的兩個細胞階段,具有pptr-2對照的N2也顯示出野生型發育,這意味著所觀察到的缺陷不僅是pptr-2功能降低的產物,并且具有L4440對照胚胎的mbk-2除了存在胞質體之外大多是正常的,胞質體是沒有細胞核的額外空細胞。見表2,圖2。

表2 線蟲胚胎發育表型檢測

圖2 胚胎發育表型觀察
3 討論
對mbk-2(dd5)修飾物的大規模RNAi篩選中,發現了四種增強子(rsa-1、pptr-1、pptr-2和rsa-2)。rsa-1、pptr-1、pptr-2均編碼PP2A的調節亞單位,rsa-2與rsa-1形成復合物[1]。PP2A參與許多重要的細胞功能,包括細胞移動性[2]、細胞骨架動力學[3]、細胞周期控制[4]、細胞生長控制、細胞增殖、細胞死亡[5]、信號通路和腫瘤抑制[6]。核心PP2A酶由支架亞單位A和催化亞單位C組成,兩者都是高度保守的[7]。PP2A底物特異性是由幾個調節亞單位之一賦予的,它與核心酶相互作用形成全酶[2]。這些調節亞單位分為四個家族:B (B55或PR55)、B (B56或PR61)、B (PR48/PR72/PR130)和B (PR93/PR110)[2]。在秀麗隱桿線蟲中,已經通過矯形學鑒定了7個PP2A調節亞單位:sur-6,B家族;pptr-1和pptr-2,B族;F47B8.3,C06G1.5,rsa-1和T22D1.5,B族[8]。
PP2A調節亞單位PPTR-1是有絲分裂期間P-顆粒的穩定性和不對稱分布所必需的[9]。在胚胎的前四個分裂階段,P顆粒的不對稱分布對于確定種系和區分種系與體細胞前體是很重要的。有絲分裂后,P顆粒在種系前體中保持穩定,但在體細胞胚粒中,P顆粒蛋白逐漸降解,P顆粒核糖核酸迅速降解[9]。PPTR-1特別適用于磷顆粒的分離,但不適用于分離PAR-1/2、MEX-5。這些種質成分從磷顆粒中自由分離。在pptr-1突變胚胎中,P顆粒和核糖核酸被平均分配到體細胞和種系卵裂球,但在這些細胞類型中表現不同。在體細胞胚中,P顆粒RNA迅速消失,P顆粒在2個細胞周期后聚集成顆粒,在原腸胚形成后消失。在種系卵裂球中,P顆粒穩定并重新組裝成顆粒;只是每個部門的規模和數量都減少了[9]。本研究顯示,在單獨抑制線蟲pprt-1表達后,對蟲卵的孵化率并無影響,而聯合抑制pprt-1和mbk-2后,蟲卵的未孵化數量增加,提示pprt-1通過調劑mbk-2發揮對發育的調節作用。
PPTR-1還通過PP2A調節秀麗新桿線蟲胰島素/IGF-1信號傳導(IIS)途徑,在幼蟲發育過程中的壽命調節、達烏爾滯育、代謝和應激反應中發揮重要作用[8]。IIS在生命周期和dauer滯育控制中是必不可少的,并且涉及由幾個絲氨酸/蘇氨酸蛋白激酶介導的磷酸化級聯。激酶將信號傳遞給胰島素樣受體DAF-2以激活PDK-1激酶,后者將信號傳遞給AKT-1、AKT-2和SGK-1[10,11]。這些激酶負調節叉頭轉錄因子-16[12]。需要PP2A活性來平衡這些激酶在胰島素抵抗途徑中的作用。PPTR-1直接抑制AKT-1的功能,這積極地促進了DAF-16的核定位和活動[8]。叉頭轉錄因子DAF-16激活與壽命、應激抗性、達斡爾族形成和脂肪儲存有關的基因轉錄[8]。
本研究的實驗結果表明pptr-2是調節mbk-2功能的最強的增強子。mbk-2的兩個主要功能是控制微管依賴過程和調節種系決定因素。為了研究由參與相互作用的兩個基因的功能喪失所產生的亞細胞表型,著重研究了pptr-2的作用。結果顯示,不論是單獨抑制pprt-2,或聯合抑制pprt-2和mbk-2,線蟲蟲卵的未孵化水平均顯著升高。特別是聯合抑制,蟲卵的未孵化率可達到76.745,提示pprt-2是調控mbk-2功能的主要增強子。從DIC結果來看,mbk-2和pptr-2的相互作用在單細胞階段沒有表現出明顯的缺陷,這意味著這種相互作用在單細胞階段沒有影響原核的遷移、紡錘體的建立或假分裂的位置。在兩細胞階段,mbk-2和pptr-2相互作用涉及顯示強烈缺陷的胚胎,這與mbk-2功能的喪失非常相似。例如:極體出現在后部,多核,胞質存在,紡錘體和胞質分裂缺陷。這些可能表明pptr-2增強子與mbk-2在相同的過程中起作用,這可能是與后驗決定子分布相同的過程。P-顆粒、OMA-1、PIE-1、POS-1和其他后向決定因素的分布需要mbk-2[13]。第一次細胞分裂后,mbk-2磷酸化OMA-1使其降解。通過將MEX-5/6分布到前部,mbk-2不對稱地定位了P-顆粒、PIE-1和位置-1[9,14]。因為PPTR-1和PPTR-2在PP2A的相同亞基調節家族中。PPTR-1對有絲分裂過程中P顆粒的穩定性和不對稱分布很重要[9,14]。PPTR-2也可能影響P顆粒和其他后決定因素。交互排斥表型,P1細胞的分裂比野生型延遲得多。這種表型先前被描述為在DNA檢查點途徑中基因功能的喪失。這可能預示著mbk-2和pptr-2對于該途徑是重要的,并且是P1細胞分裂時間所必需的。
