半干旱黃土丘陵區(qū)微地形變化對人工檸條林草本分布的影響
2021-04-08 10:50:36王子婷柴春山張洋東劉冬皓
草地學(xué)報 2021年2期
王子婷, 楊 磊, 李 廣, 柴春山, 張洋東, 劉冬皓
(1.甘肅農(nóng)業(yè)大學(xué)林學(xué)院, 甘肅 蘭州 730070; 2.甘肅省林業(yè)科學(xué)研究院, 甘肅 蘭州 730020;3.中國科學(xué)院生態(tài)環(huán)境研究中心城市與區(qū)域生態(tài)國家重點實驗室, 北京 100085)
黃土高原丘陵溝壑區(qū)是我國生態(tài)環(huán)境脆弱、水土流失嚴重的區(qū)域之一,也是我國西部生態(tài)安全屏障的重要組成部分[1-3]。隴中黃土丘陵溝壑區(qū)地處甘肅省中部,屬于黃土高原丘陵溝壑區(qū)第Ⅴ副區(qū),梁峁坡主要分布在15°~25°區(qū)間,水土流失面積占本區(qū)總面積的96.8%[4-6]。氣候干旱和人類活動的長期干擾導(dǎo)致該區(qū)一直是我國植被恢復(fù)建設(shè)的重點關(guān)注區(qū)域[7]。該區(qū)早在20世紀50年代就開展了一系列小流域綜合治理工作,90年代末又實施了退耕還林(草)等生態(tài)恢復(fù)工程[8]。坡面建植的人工林草植被有效抑制了水土流失,減少了土壤侵蝕,提高了土壤碳氮儲量,改善了區(qū)域生態(tài)環(huán)境[2,9-10]。然而,人工植被恢復(fù)會過度消耗坡面土壤水分,降低林下草本植物的多樣性,出現(xiàn)植被退化等現(xiàn)象,從而影響植被群落的穩(wěn)定[11]。因而,有必要針對坡面微地形變化開展人工植被恢復(fù)對草本多樣性影響的研究,確定地形、植被和土壤水分的互饋關(guān)系,為人工林地的管理及穩(wěn)定性研究提供數(shù)據(jù)支撐和理論依據(jù)。
黃土丘陵區(qū)坡面地形變化改變降雨入滲過程引起土壤水分及養(yǎng)分的再分配,進而影響林木和草本植物的分布格局。坡向及坡位變化影響水熱組合條件,從而決定了林木和草本的生長、密度或蓋度,這在人工植被恢復(fù)后期表現(xiàn)尤為明顯[12-13]。在半干旱黃土區(qū)坡面植被恢復(fù)過程中,依據(jù)坡度變化利用工程整地措施(如魚鱗坑、水平階和反坡臺地)匯集降雨,從而減少水土流失,提高土壤水分的有效性,最終促進灌木和草本植物生長及更新[14-17]。隴中黃土區(qū)坡面微地形改造栽植灌木或喬木,在一定程度上提高了水土保持、固碳等生態(tài)服務(wù)功能,但也導(dǎo)致土壤水分呈現(xiàn)不同程度的匱缺[18]。同樣,微地形改變坡面土壤水分及養(yǎng)分,也會影響草本的分布及多樣性。汝海麗等[19]研究發(fā)現(xiàn),黃土丘陵區(qū)陰坡草本生物量及多樣性大于陽坡,下坡位草本生物量及多樣性大于中、上坡位;王子婷等[13]研究發(fā)現(xiàn),坡向和坡位等微地形決定人工苜蓿草地的退化程度,苜蓿(MedicagosativaL.)退化導(dǎo)致人工草地向天然草地轉(zhuǎn)變。在人工檸條(Caraganakorshinskii)林地恢復(fù)過程中,林地密度及恢復(fù)年限等均會影響草本群落分布及多樣性[20-21]。因此,坡面采用水平階整地方式栽植檸條,檸條的生長及微地形變化會影響草本分布及多樣性。鑒于此,本研究在甘肅省定西市安定區(qū)龍灘小流域選擇成熟的檸條林地為研究對象,解析微地形改變及栽植檸條對水平階內(nèi)及階間坡面草本分布及多樣性的影響,確定人工檸條林地形變化和土壤水分對坡面草本分布及多樣性的影響。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于甘肅省定西市安定區(qū)巉口鎮(zhèn)龍灘小流域(104°27′~104°32′ E,35°43′~35°46′ N),屬典型半干旱黃土丘陵區(qū),流域面積16 km2,平均海拔1 900 m。研究區(qū)年平均氣溫6.8℃,1月份平均氣溫—7.9℃,極端最高溫38.5℃,平均無霜期152 d,平均日照時數(shù)2 052 h。該區(qū)多年平均降水量為386 mm,降雨主要集中在7—9月,潛在蒸發(fā)量1 439 mm,年平均相對濕度72%,干燥度1.9。流域內(nèi)土壤以黃綿土為主,有機質(zhì)含量低,土壤貧瘠且易侵蝕。天然植被以多年生草本為主,主要草本種類有長芒草(Stipabungeana)、賴草(Leymussecalinus)和阿爾泰狗娃花(Heteropappusaltaicus)等,人工種植的灌木及喬木樹種有檸條、側(cè)柏(Platycladusorientalis)和油松(Pinustabuliformis)等,其中人工檸條林地面積較大且分布較廣。
1.2 研究方法
1.2.1監(jiān)測樣點布設(shè) 在研究區(qū)內(nèi)選擇一個全部種植檸條灌木的典型坡面,該坡面檸條從坡頂至坡底沿等高線以水平階整地的造林方式種植于1984年。水平階寬度為1~1.5 m,檸條均栽植于水平階內(nèi)中部位置,兩水平階的階間坡面寬約3~5 m。沿坡面自上而下選擇上、中和下3個坡位,在每個坡位上選擇水平階內(nèi)和階間坡面2種微生境。上、中和下坡位階間坡面的坡度分別為25°,33°和22°,水平階整地后階內(nèi)坡度接近零度。水平階之間為自然坡面,植被以草本為主,人為擾動較小。
1.2.2數(shù)據(jù)采集 于2019年8月在上、中和下坡位分別選取4個地點,每個地點包括水平階內(nèi)和階間坡面2種微生境,每種微生境選擇50 cm×50 cm范圍調(diào)查草本群落組成及生物量,在每個草本樣方附近10 m×10 m范圍內(nèi)每木調(diào)查檸條生長情況。利用GPS記錄各監(jiān)測樣點的經(jīng)緯度及海拔信息,利用坡度儀和羅盤分別測定坡度和坡向。統(tǒng)計樣方內(nèi)草本物種數(shù)、高度及蓋度,隨后將草本地上部分進行刈割,樣品帶回實驗室稱重后在105℃下殺青1 h后再在80℃下烘干72 h,分別獲得草本地上生物量及其含水量。不同坡位檸條的生長狀況不同,中坡位檸條密度(3 000 株·ha-1)高于上、下坡位(1 900和2 200 株·ha-1),下坡位檸條株高、平均冠幅及基徑(2.25 m,2.73 m和13.27 mm)均高于中、上坡位(1.42 m,1.64 m和10.16 mm;1.70 m,2.36 m和9.56 mm)。利用環(huán)刀法測定0~20 cm土壤水分含量和容重。上、中坡位土壤水分略高于下坡位,中坡位水平階內(nèi)土壤水分高于階間坡面,上坡位和下坡位2種微生境內(nèi)土壤水分含量相差較小(圖1)。
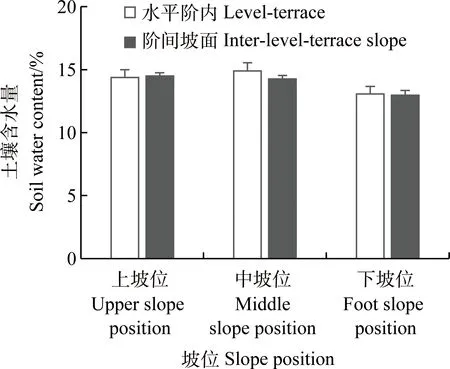
圖1 上、中和下坡位不同微生境內(nèi)土壤含水量變化
1.2.3數(shù)據(jù)統(tǒng)計 統(tǒng)計各監(jiān)測樣點每個樣方內(nèi)草本物種數(shù),使用每種草本的相對高度和相對蓋度計算其重要值(IV)[22]。利用重要值計算草本的相對多度,然后按照豆科、禾本科、菊科、薔薇科和其它科分別統(tǒng)計上、中和下坡位不同微生境下草本物種數(shù),計算每一類草本物種數(shù)占總物種數(shù)的比重。利用二因素方差分析確定坡位和微地形(水平階內(nèi)和階間坡面)對不同草本類群(科)、主要草本種及地上生物量的影響,使用配對t檢驗確定2種微生境之間的差異,統(tǒng)計分析使用SPSS 21.0軟件包。
采用非度量多維測度(Non-metric multidimensional scaling,NMDS)和相似性分析(Analysis of similarities,ANOSIM)分析坡面及坡面不同部位水平階內(nèi)和階間坡面草本群落的相似性,SIMPER分析確定坡面及不同坡位水平階內(nèi)和階間坡面草本群落的相異性及主要草本物種的貢獻率,分析使用Past 4.01。利用方差分析確定坡位(上、中和下)和微生境(水平階內(nèi)和階間坡面)對草本群落物種數(shù)、主要草本種、生物量及含水量的影響。采用RDA分析坡面草本群落分布與地形變化的關(guān)系,確定坡位、微生境及土壤環(huán)境對草本群落分布的貢獻率,分析使用Canaco 5.0。利用Pearson分析研究草本群落結(jié)構(gòu)和主要草本種與草本生物量、土壤含水量及坡位的相關(guān)性,統(tǒng)計分析使用SPSS 21.0。
2 結(jié)果與分析
2.1 不同坡位水平階內(nèi)和階間坡面草本群落組成
在水平階內(nèi)和階間坡面2種微生境下的24個草本樣方內(nèi)共確定12科、23屬和28種草本植物。其中以禾本科(占總物種數(shù)的7.1%)、豆科(14.3%)、菊科(32.1%)和薔薇科(14.3%)為主要類群,其它草本占32.2%;多年生草本占85.7%,一年生草本僅占14.3%(表1)。禾本科長芒草、賴草,薔薇科伏毛山莓草(Sibbaldiaadpressa)和菊科阿爾泰狗娃花、茵陳蒿(Artemisiacapillaris)、冷蒿(Artemisiafrigid)及束傘亞菊(Ajaniaparviflora)是人工檸條林地主要草本種,它們對坡位及微地形變化的響應(yīng)模式不同。長芒草主要分布在下坡位,其在下坡位的重要值顯著高于中、上坡位(P<0.05);束傘亞菊的變化趨勢與之相反,其在中、上坡位的重要值顯著高于下坡位(P<0.05)。同時,研究還發(fā)現(xiàn)草本種對微地形的響應(yīng)還隨坡位的不同而異,長芒草和束傘亞菊主要分布在階間坡面,長芒草在中坡位、束傘亞菊在上坡位階間坡面的重要值顯著高于水平階內(nèi)(P<0.05);茵陳蒿的重要值變化趨勢與長芒草和束傘亞菊相反,主要分布在水平階內(nèi)。
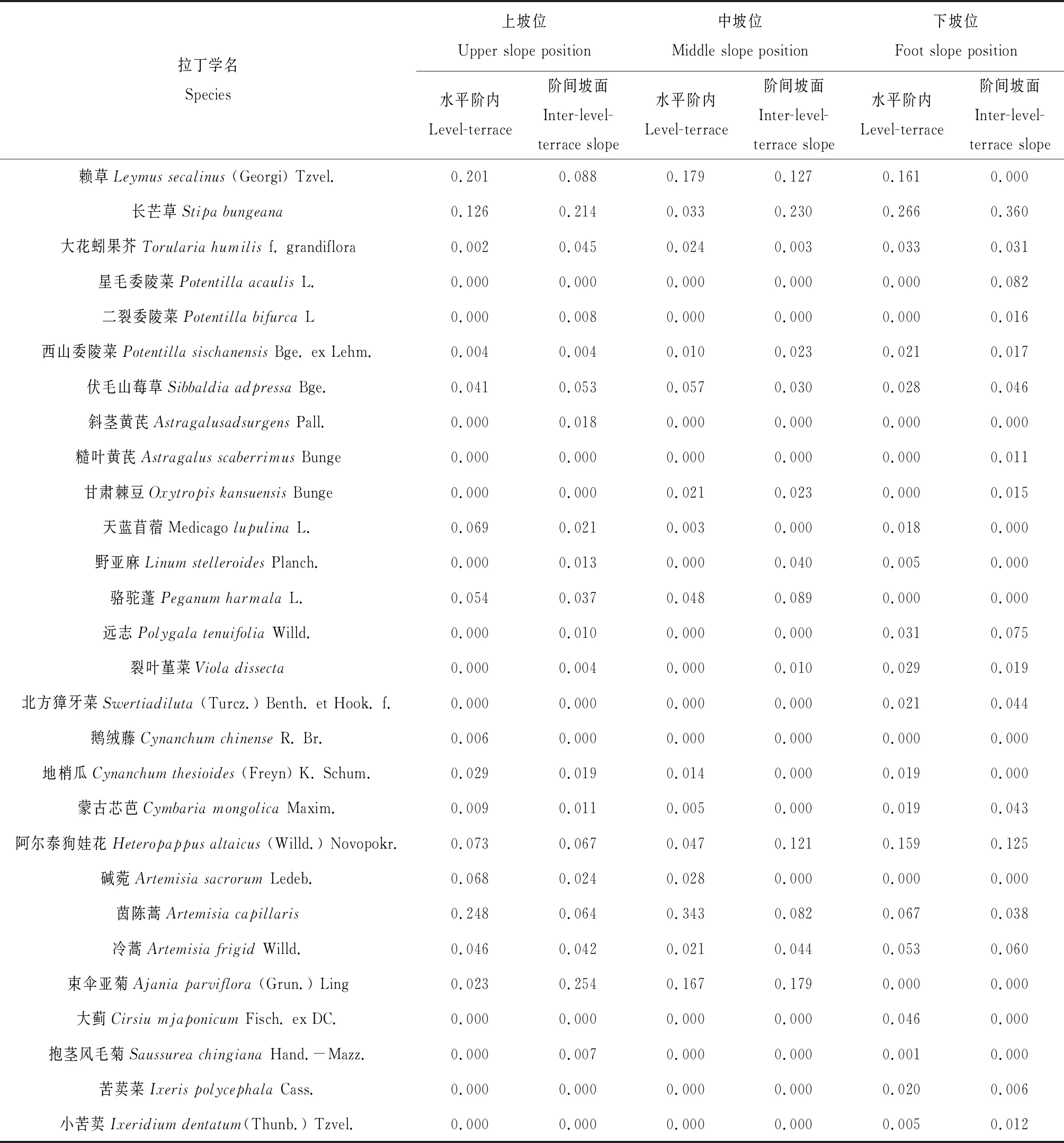
表1 不同坡位水平階內(nèi)和階間坡面草本的重要值
不同種類的草本對微生境(水平階內(nèi)和階間坡面)的響應(yīng)模式隨坡位的不同而不同,禾本科、豆科、菊科、薔薇科和其它草本物種數(shù)及相對多度在上、中坡位的2種微生境下相差較小,而在下坡位存在明顯變化(圖1)。水平階內(nèi)禾本科、菊科和其它草本物種數(shù)及相對多度均高于階間坡面,其中禾本科物種數(shù)及相對多度在水平階內(nèi)顯著高于階間坡面(圖1)。豆科和薔薇科草本變化趨勢與禾本科、菊科和其它草本相反,其中薔薇科草本物種數(shù)及相對多度在水平階內(nèi)顯著低于階間坡面(圖2)。
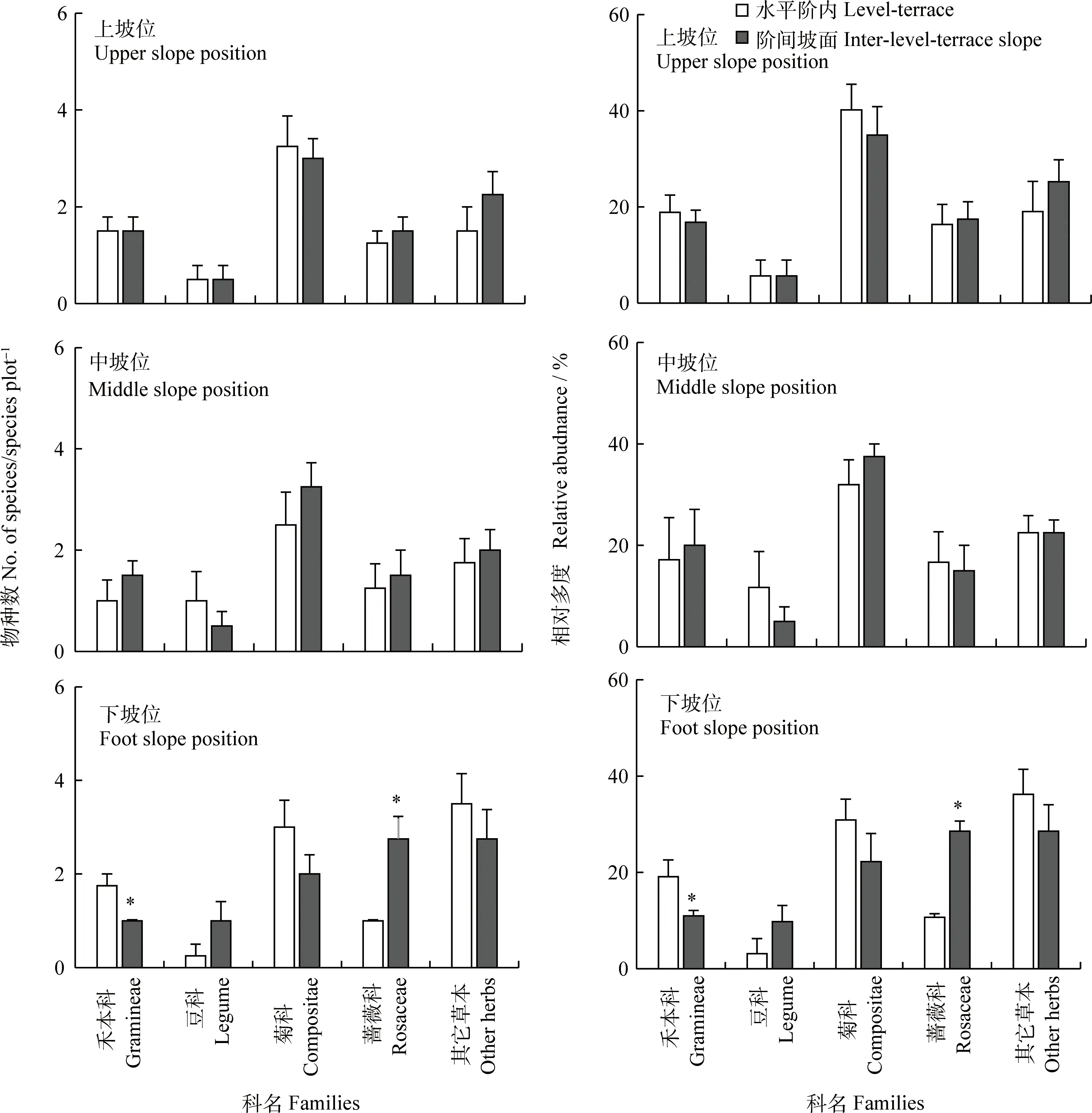
圖2 上、中和下坡位水平階內(nèi)和坡面5類草本物種數(shù)和相對多度變化
2.2 不同坡位水平階內(nèi)和階間坡面草本群落相似性比較
坡面上、中和下坡位水平階內(nèi)和階間坡面草本群落的NMDS排序結(jié)果表明,第一軸解釋了55.1%的群落變異,第二軸解釋了10.7%的群落變異,水平階內(nèi)和階間坡面草本群落組成明顯不同并隨坡位不同而異(圖3)。ANOSIM分析結(jié)果表明,水平階內(nèi)和坡面之間草本群落組成明顯不同(R2=0.18,P=0.011)。上、中和下坡位水平階內(nèi)和階間坡面草本群落組成存在一定差異(ANOSIM:R2=0.39,P=0.055;R2=0.47,P=0.057;R2=0.32,P=0.057)。
SIMPER分析表明,不考慮坡位的影響,水平階內(nèi)和階間坡面草本群落的相異性為65.9%,其中冷蒿、茵陳蒿、長芒草、賴草、束傘亞菊和阿爾泰狗娃花在2種微生境內(nèi)分布不同,它們貢獻了2種微生境草本群落差異的68.4%。上坡位水平階內(nèi)和階間坡面草本群落SIMPER分析結(jié)果表明,二者之間的相異性為75.2%,冷蒿、束傘亞菊、賴草、茵陳蒿和長芒草在2種微生境內(nèi)分布不同,為上坡位水平階內(nèi)和階間坡面草本群落差異貢獻了74.1%。中坡位水平階內(nèi)和階間坡面草本群落SIMPER分析結(jié)果表明,二者之間的相異性為60.0%,茵陳蒿、長芒草、賴草、束傘亞菊、阿爾泰狗娃花和駱駝蓬(Peganumharmala)在2種微生境內(nèi)分布不同,它們貢獻了2種微生境草本群落差異的77.4%。下坡位水平階內(nèi)和階間坡面草本群落SIMPER分析結(jié)果表明,二者之間的相異性為50.7%,賴草、長芒草、冷蒿、星毛委陵菜(Potentillaacaulis)、遠志(Polygalatenuifolia)、茵陳蒿和阿爾泰狗娃花在2種生境內(nèi)分布不同,它們貢獻了2種微生境草本群落差異的63.3%。
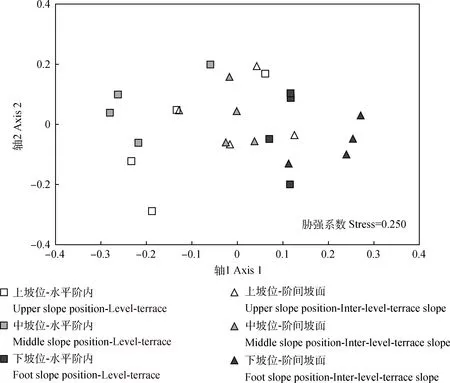
圖3 上、中和下坡位水平階內(nèi)和階間坡面草本群落的NMDS排序圖
2.3 不同坡位水平階內(nèi)和階間坡面草本生物量比較
坡位和微生境對草本地上生物量及含水量影響的二因素方差分析結(jié)果表明,坡位和微生境顯著影響坡面草本生物量(P<0.05),而兩者對草本生物量的交互影響不顯著。上、中坡位水平階內(nèi)草本生物量顯著高于階間坡面(P<0.05),下坡位水平階內(nèi)草本生物量略高于階間坡面,上、中和下坡位水平階內(nèi)草本生物量分別是坡面生境的2.2,1.9和1.3倍(圖4)。坡面草本含水量與生物量變化不同,微生境對草本含水量的影響顯著(P<0.05),而坡位和微生境對草本含水量的交互影響不顯著。上、中和下坡位水平階內(nèi)草本含水量均顯著高于坡面生境,其草本含水量分別是坡面生境的1.12,1.11和1.15倍(圖4)。
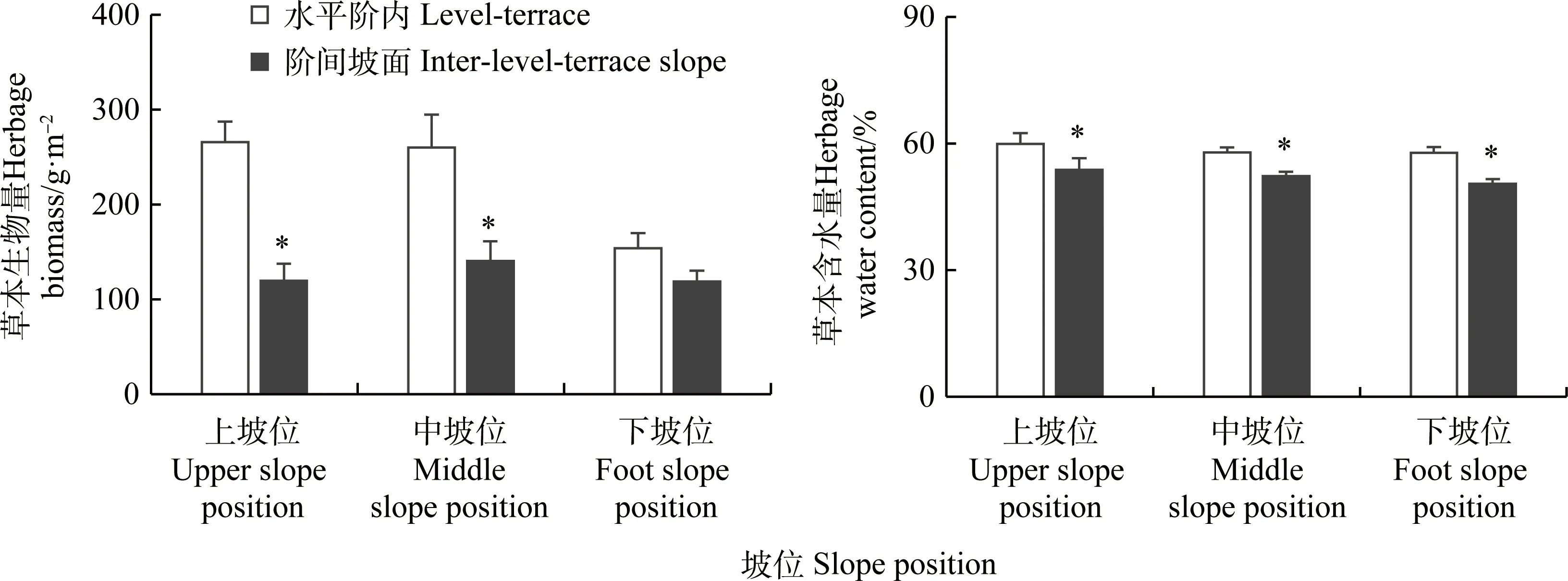
圖4 上、中和下坡位水平階內(nèi)和階間坡面草本生物量和含水量變化
2.4 地形、土壤和植被等環(huán)境要素對草本群落分布的影響
利用RDA和pRDA分析不同坡位水平階內(nèi)及階間坡面草本群落組成與4個環(huán)境因子(坡位、水平階內(nèi)及階間坡面、土壤水分和容重)的關(guān)系,確定影響坡面草本分布的關(guān)鍵環(huán)境因子。坡面草本與環(huán)境因子的排序結(jié)果表明,第1軸解釋了23.7%的草本群落變化(Eigenvalue=0.237,F(xiàn)=6.10,P<0.001),所有的排序軸解釋了38.9%的草本群落變化(Eigenvalue=0.389,F(xiàn)=3.00,P<0.001),排序結(jié)果較好的反映了地形和土壤水分對草本分布的影響。pRDA分析結(jié)果表明,土壤含水量對坡面草本分布影響最大(解釋了16.8%的變異),其次是微生境(即水平階內(nèi)及階間坡面)(12.9%)和坡位(6.1%),3個因子累積解釋了35.8%坡面草本群落變異(圖5)。
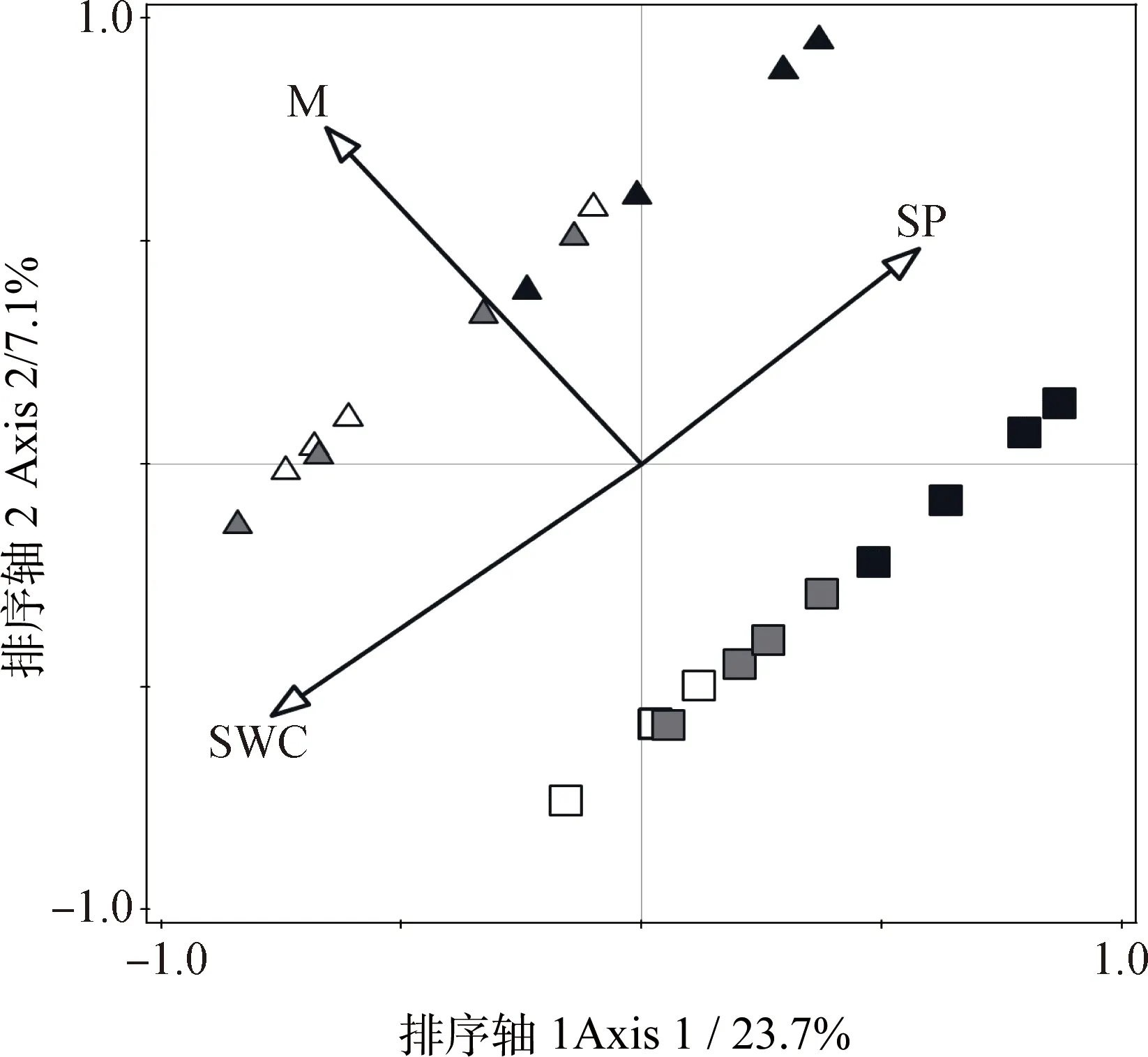
圖5 坡面草本群落分布與環(huán)境因子的RDA排序圖
土壤水分與草本地上生物量及物種豐富度Pearson相關(guān)分析結(jié)果表明,土壤水分與草本地上生物量及物種豐富度的相關(guān)性均較小(r=0.27,P=0.201;r=-0.10,P=0.637),草本生物量與草本物種數(shù)呈顯著負相關(guān)(r=-0.42,P=0.042)。此外,研究還發(fā)現(xiàn),階間坡面土壤水分與草本生物量的相關(guān)性較小(r=-0.01,P=0.980),而水平階內(nèi)草本地上生物量與土壤水分呈正相關(guān)(r=0.51,P=0.090)。草本生物量與主要草本種Pearson相關(guān)分析表明,長芒草和阿爾泰狗娃花與草本生物量呈顯著負相關(guān),與駱駝蓬和茵陳蒿呈顯著正相關(guān)(表2)。土壤含水量與主要草本種Pearson相關(guān)分析表明,長芒草與土壤含水量呈顯著負相關(guān),與茵陳蒿和束傘亞菊呈顯著正相關(guān)(表2)。此外,研究還發(fā)現(xiàn)水平階內(nèi)主要草本種與坡位的相關(guān)性較小,階間坡面束傘亞菊與坡位呈顯著負相關(guān),它主要分布在中、上坡位;長芒草與坡位呈顯著正相關(guān),它主要分布在中、上坡位(表2)。
3 討論
黃土高原丘陵區(qū)人工恢復(fù)檸條,檸條生長通過改變表層土壤水分及養(yǎng)分變化驅(qū)動草本群落演替過程[20,23]。坡面微地形變化改變了人工植被的生長狀況或退化程度,進而影響草本群落結(jié)構(gòu)及生物量。前期的研究已經(jīng)發(fā)現(xiàn),坡面下坡位人工檸條生長好于中、上坡位,下坡位苜蓿草地的退化程度也較中、上坡位低,從而影響了不同坡位草本群落的恢復(fù)演替過程[12-13]。本研究發(fā)現(xiàn),人工檸條林地下坡位草本地上生物量低于中、上坡位,而水平階內(nèi)草本的生物量明顯高于階間坡面,且在上、中坡位表現(xiàn)尤為明顯,這與周萍等[24]和汝海麗等[19]在陜北黃土丘陵區(qū)坡面草本生物量及多樣性分布規(guī)律的研究結(jié)果不同。水平階整地及檸條生長改變坡面草本地上生物量的分布格局,中、上坡位水平階內(nèi)草本地上生物量高于下坡位,這可能與檸條生長狀況和表層土壤水分的改善有關(guān)[25-26]。坡面水平階整地栽植檸條,微地形限制了中、上坡位檸條生長,這也提高了表層土壤水分含量(中、上坡位表層土壤含水量略高于下坡位),促進了一些草本種生長,從而提高了中、上坡位草本的生物量,這與程積民等[14]和郭璞等[27]在內(nèi)蒙荒漠化草原的研究結(jié)果相近。坡面水平階整地會提高土壤水分有效性,從而提高草本的地上生物量;栽植檸條可以提高荒漠化生境草本的生物量,木本和草本植物生物量的增加也會降低草本多樣性[14,27]。此外,研究還發(fā)現(xiàn)坡面檸條林地草本生物量與物種豐富度呈顯著負相關(guān),草本生物量和草本多樣性對地形變化的響應(yīng)不一致。水平階整地栽植檸條并沒有改變坡面草本物種豐富度的分布規(guī)律,坡面草本分布規(guī)律與寇萌等[28]在黃土丘陵區(qū)的研究結(jié)果相近,而與王子婷等[13]關(guān)于坡面退化苜蓿草地的研究結(jié)果不同,說明水平階整地栽植檸條在一定程度上降低了坡位變化對草本分布及多樣性的影響。由此可見,坡面人工恢復(fù)灌木的生長狀況、栽植密度及退化程度等影響剖面土壤水分含量,進而影響林下草本的生物量及多樣性,驅(qū)動林下草本群落的恢復(fù)演替過程[21,29]。因而,黃土丘陵區(qū)植被管理需要依據(jù)坡面立地條件制定天然和人工植被管理策略,這樣才能更好維持植被的多樣性與穩(wěn)定性[30-31]。
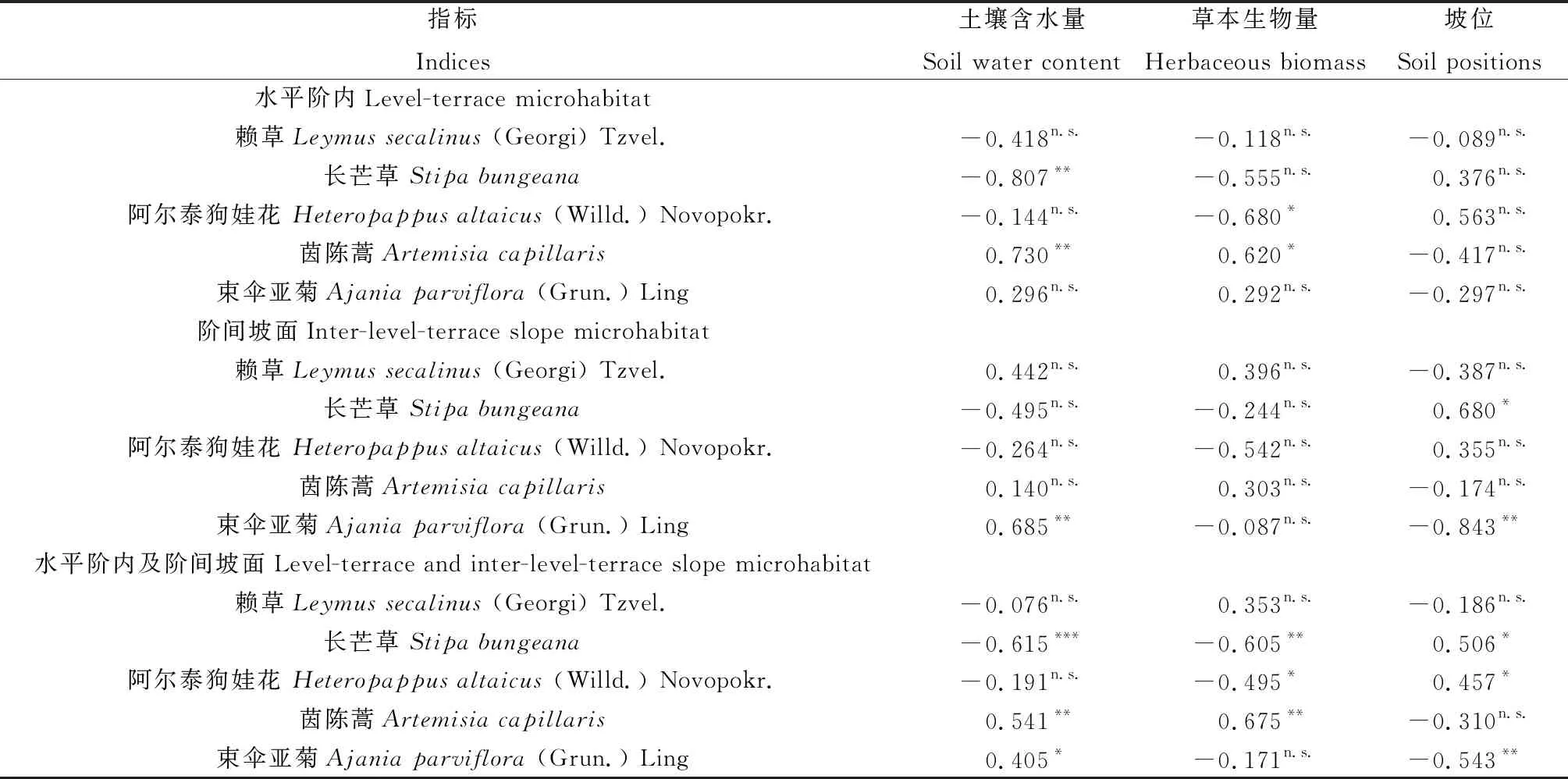
表2 水平階內(nèi)和階間坡面主要草本種與土壤水分、草本生物量和坡位的Pearson和Spearman相關(guān)系數(shù)
上、中和下坡位水平階內(nèi)與階間坡面草本群落組成明顯不同,下坡位草本群落的相異性低于中、上坡位,下坡位禾本科和薔薇科草本物種數(shù)及相對多度明顯不同,這與下坡位水平階內(nèi)及階間坡面草本生物量低于中、上坡位有關(guān)。研究區(qū)下坡位檸條密度略低于中、上坡位,其生長好于中、上坡位,而表層土壤水分變化相反,這也導(dǎo)致中、上坡位茵陳蒿和束傘亞菊的重要值及生長好于下坡位,從而提升中、上坡位水平階內(nèi)草本的地上生物量。樊如月等[21]和劉燕萍等[29]研究發(fā)現(xiàn),草本種類數(shù)隨著檸條林分密度的增加而降低且旱生植物比例增大,隔行平茬檸條林地與對照相比提高了草本地上生物量,但過低的林地密度又降低了草本的地上生物量,這說明適度降低成熟檸條林地密度可以促進草本群落恢復(fù)。此外,本研究還發(fā)現(xiàn)一些非禾本科草本重要值及生物量的增加也限制長芒草等禾本科和菊科等一些地帶性草本的生長,從而降低了中、上坡位草本的多樣性。由此可見,水平階整地栽植檸條,微地形變化不但影響檸條的生長,它還與灌木一起通過調(diào)節(jié)水熱組合條件影響草本群落組成及多樣性。坡面草本群落分布與坡位、微地形和土壤環(huán)境的定量分析結(jié)果表明,影響坡面草本群落結(jié)構(gòu)的主要環(huán)境因子是土壤水分、微生境和坡位,這與汝海麗等[19]、陳文思等[30]和梁香寒等[23]的研究結(jié)果相近,而與李冬梅等[32]的研究結(jié)果略有不同。表層土壤水分與草本生物量及物種豐富度的相關(guān)性較小,物種豐富度與草本生物量呈顯著負相關(guān),這與自然草地生態(tài)系統(tǒng)的研究結(jié)果不同等[33]。然而,本研究還發(fā)現(xiàn)水平階內(nèi)和階間坡面土壤水分與草本地上生物量的關(guān)系不同,水平階內(nèi)草本生物量與土壤水分呈正相關(guān),而在階間坡面二者的相關(guān)性則較小。坡面中、上坡位較高的表層土壤水分含量提高了水平階內(nèi)草本地上生物量,而下坡位隨著檸條生長導(dǎo)致表層土壤水分較低,這也降低了草本的地上生物量。以上研究結(jié)果與劉艷萍等[29]關(guān)于平茬檸條林地的研究結(jié)果略有不同,平茬檸條林地草本植物多樣性各指數(shù)及生物量均與土壤剖面含水量呈負相關(guān)關(guān)系,其中物種豐富度及多樣性與表層土壤水分呈顯著負相關(guān),這說明降低成熟檸條林地密度、提高表層土壤水分含量有利于恢復(fù)草本的多樣性[21]。賈希洋等[34]在寧夏荒漠草原區(qū)研究也發(fā)現(xiàn),隔兩帶平茬一帶處理下植被物種總數(shù)、密度、高度和地上生物量和土壤含水量最高,物種豐富度和多樣性在隔兩帶平茬一帶和隔一帶平茬一帶處理下均較高,這說明不同地域條件下平茬處理對草本分布的影響略有不同。成熟人工檸條林地坡面微地形改變通過調(diào)節(jié)檸條的生長還影響了草本群落組成,它還通過改變坡面草本生物量分布格局來影響草本群落組成。茵陳蒿等多年生草本決定草本地上生物量,它們也通過競爭限制了長芒草等禾本科草本植物的生長,從而降低了中、上坡位草本的物種豐富度,而下坡位隨著草本生物量的降低,草本的物種豐富度又略有增加。總之,利用水平階改造坡面栽植檸條,隨著檸條蓋度及生物量的增加,它對林下草本多樣性的抑制作用在增強,這種影響因坡位不同而異。此外,本研究還發(fā)現(xiàn),坡位改變了人工檸條林地草本植物地上生物量的分布格局,隨著草本地上生物量的降低物種豐富度呈增加的趨勢,而長芒草等地帶性草本的比重也隨之增加。由此可見,坡面微地形改造恢復(fù)灌木均會影響土壤水分,中、上坡木本植物生長較差,而一些菊科的多年生草本生長大幅提升地上生物量,抑制了禾本科等地帶性草本的生長,結(jié)合地形變化進行檸條林地的疏化管理則可降低土壤水分消耗,提高林下地帶性草本的多樣性[35-36]。
4 結(jié)論
半干旱黃土丘陵區(qū)坡面人工檸條林地草本分布及多樣性明顯受微地形變化的影響,水平階內(nèi)草本生物量及含水量均高于階間坡面,不同坡位水平階內(nèi)和階間坡面草本組成的相異性為上坡位(75.2%)>中坡位(60.0%)>下坡位(50.7%)。pRDA排序表明,土壤含水量、坡位和微生境是影響坡面草本分布的主要環(huán)境因子,解釋了草本群落35.8%的變異。相關(guān)分析表明,土壤含水量與長芒草的重要值呈顯著負相關(guān),與茵陳蒿和束傘亞菊的重要值呈顯著正相關(guān)。由此可見,因下坡位檸條生長好于中、上坡位,從而降低了表層土壤水分含量,二者相互作用限制了下坡位茵陳蒿和束傘亞菊等多年生草本生長,促進了長芒草等地帶性草本的恢復(fù),因此成熟檸條林地的疏化管理需要考慮微地形變化,這樣才能更好地緩解土壤水分匱缺并維持草本多樣性。
