光強對4種鴨跖草科植物葉片生理特性和超微結構的影響
2021-04-08 10:55:36李子葳楊宇佳楊小梅
草地學報 2021年2期
關鍵詞:植物
陳 斌, 薛 晴, 李子葳, 楊宇佳, 楊小梅, 薄 杉, 李 強, 何 淼
(東北林業大學園林學院, 黑龍江 哈爾濱 150040)

紫鴨跖草(Commelinapurpurea)、‘花葉’水竹草(Tradescantiaflurnuensis‘Variegata’)、吊竹梅(Tradescantiazebrina)、‘綠葉’水竹草(Tradescantiaflurnuensis‘Vairidia’)是鴨跖草科(Commelinaceae)的4種植物,植株低矮、葉色豐富、耐瘠薄、莖多呈匍匐狀、莖節處易生氣生根、繁殖能力強、成坪覆蓋速度快且價格低廉,現已經作為地被植物在園林綠化中應用,具有較強的光適應能力,但每種植物的適光策略目前尚不清晰。因此,本試驗通過人工模擬不同立地條件下的光強,通過遮光網設置5種光環境對紫鴨跖草、‘花葉’水竹草、吊竹梅和‘綠葉’水竹草進行光處理,測定不同光強下4種植物的生理生化指標和葉肉細胞的超微結構,揭示4種植物的適光策略和潛能,以期為其在園林中栽培應用和進一步大范圍的推廣提供科學依據。
1 材料和方法
1.1 試驗材料
試驗于2019年3—7月在黑龍江省哈爾濱市東北林業大學花卉研究所苗圃進行。以紫鴨跖草、‘花葉’水竹草、吊竹梅、‘綠葉’水竹草為材料(表1),3月初選取當年生健壯的枝條,剪取頂部長約5 cm的莖段作為插條,基質采用75%腐殖土+25%蛭石混合,經120℃高溫滅菌后裝入直徑12 cm高18 cm的花盆中,采用浸盆法充分浸潤盆中的栽培基質,然后進行扦插。每盆扦插2個莖段,每種試驗材料各60盆。扦插完畢后放置在育苗室內緩苗,緩苗期間的環境條件為:溫度24~26℃,相對濕度60%~70%,12 h光照,白天光強為480 μmol·m-2·s-1。緩苗1個月后,選取長勢旺盛,無病蟲害的植株進行試驗。

表1 植物材料
1.2 試驗方法
于2019年4月30在苗圃內利用不同密度的黑色尼龍遮光網設置5種光強梯度(分別為自然光強的100%,75%,50%,25%,5%),每個處理間隔50 cm,頂部遮光網距植株體頂端30 cm,每個光強梯度每種植物材料各處理12盆,3個重復。每隔5 d澆水一次,處理45 d后,進行各項指標的測定。
1.3 指標測定
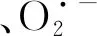
二氨基聯苯胺法(diaminobenzidine,DAB)染色測定H2O2:摘取莖頂部第3~5片成熟完整的葉片,放入pH為5.8的DAB染色液(1 mg·mL-1)中,在28℃下過夜避光保存,然后放入80%的乙醇中水浴煮沸若干次,直至葉片顏色完全脫去,并用相機拍照。
葉片超微結構的觀察:取健壯植株的第3片功能葉,在葉脈兩側各取1 mm×3 mm的矩形小塊,每組處理取3片。用濃度為2.5%的戊二醛溶液和1%的鋨酸進行雙固定,固定后用pH6.8的磷酸緩沖液漂洗;然后用不同濃度的乙醇及丙酮溶液依次脫水,并用環氧樹脂包埋;采用LKB-5型超薄切片機進行切片,切片經醋酸鈾-枸櫞酸鉛雙染色后,于H7650型透射電子顯微鏡下對材料進行觀察并拍照[17],每個處理取10個視野,統計每個視野中每個葉肉細胞內的葉綠體、線粒體、淀粉粒數量,并在每個視野中隨機選取5個葉綠體,統計每個葉綠體中嗜鋨顆粒數量。
1.4 數據整理與分析
用Excel 2016對數據進行整理和作圖,利用SPSS 22.0對數據進行方差分析,用Duncan法進行多重比較(P<0.05),并對處理結果進行雙因素(ANOVAs)分析。參照李京蓉等[18]的方法對4種植物采用隸屬函數法進行綜合評價。
2 結果與分析
2.1 光強對滲透調節物含量的影響
不同的光強對4種鴨跖草科植物的SS含量影響不同(圖1)。CP的SS含量在L0~L3光強間無顯著差異,而在L4光強下顯著降低(P<0.05),TF的SS含量隨光強的降低顯著降低(P<0.05),與L0相比,在L4條件下降低了58.91%,而TV的SS含量在L0~L2光強間無顯著差異,但均顯著高于L3和L4條件。TZ的SS含量隨光強的降低呈先升高后降低的趨勢,在L3條件下上升到最高值,在L4條件下降到最低值,與最高值相比顯著降低了22.06%(P<0.05);
在不同的光強下,4種植物的SP含量呈相同的變化趨勢(圖1),均隨光強的降低而顯著降低(P<0.05)。與L0條件相比,CP,TF,TZ和TV在L4條件的SP含量分別降低了48.68%,65.31%,70.28%,57.39%,其中TZ的降低幅度最大。
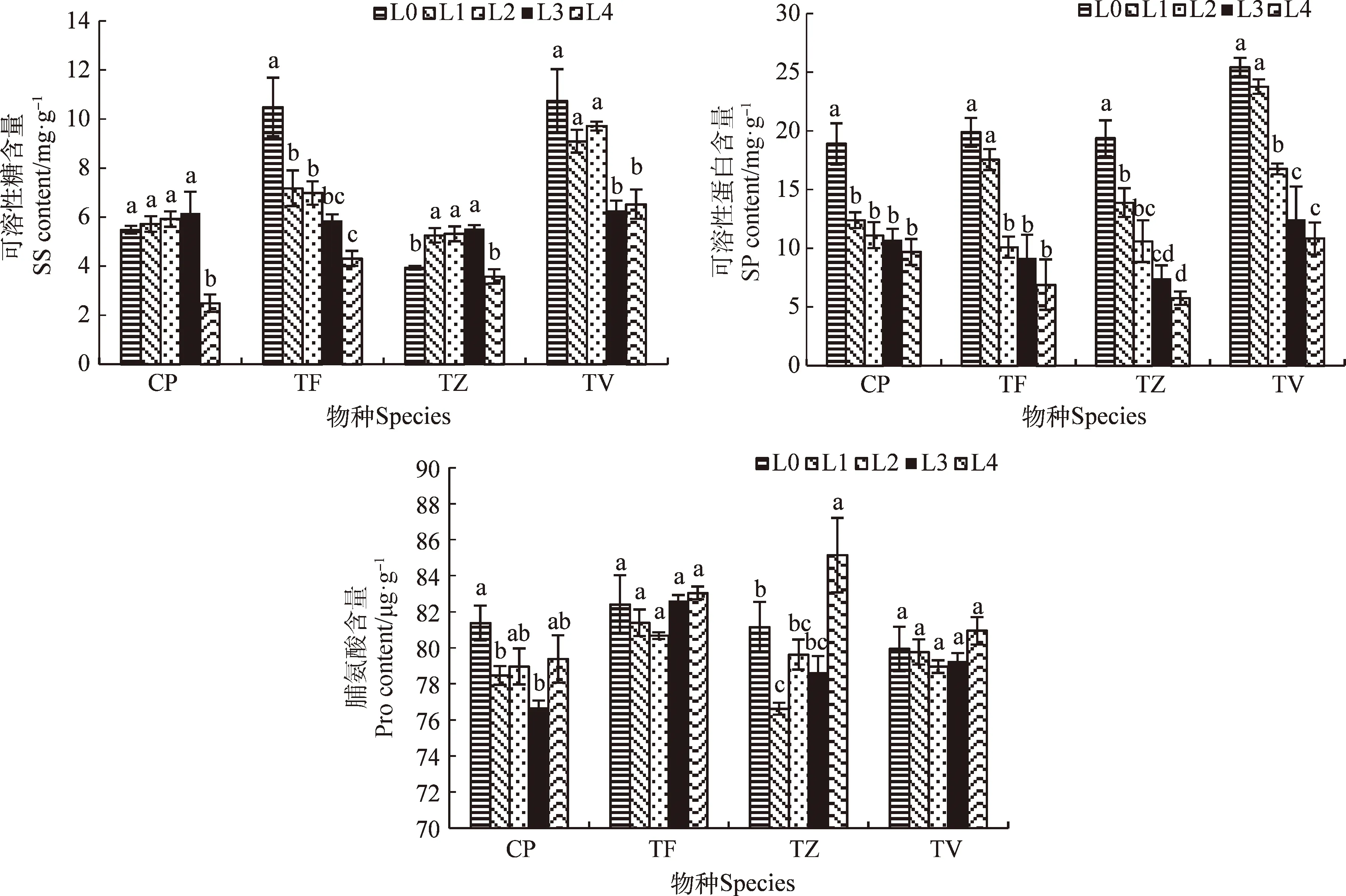
圖1 不同光強對滲透調節物含量的影響
4種植物的Pro含量隨光強的降低呈先降低后增加的趨勢,且在L0和L4條件下的Pro含量均高于L1,L2和L3光強條件,但TF和TV在各處理間的差異均不顯著(圖1)。CP在L0條件下達到峰值,是最低值(L3條件下)的1.06倍,TZ在L4條件下達到峰值,是最低值(L1條件下)的1.11倍。
2.2 光強對MDA含量的影響
不同的光強下,4種植物的MDA含量的變化趨勢相同,均隨光強的降低而顯著降低(P<0.05)(圖2)。與L0相比,CP,TF,TZ和TV在L4條件的MDA含量分別降低了42.60%,36.87%,73.65%,43.44%,且TZ和TV在L0~L3條件下的MDA含量高于CP和TF,這表明強光條件導致4種植物產生了膜脂過氧化反應,且TZ和TV的膜系統損傷更大。
2.3 光強對H2O2和含量的影響
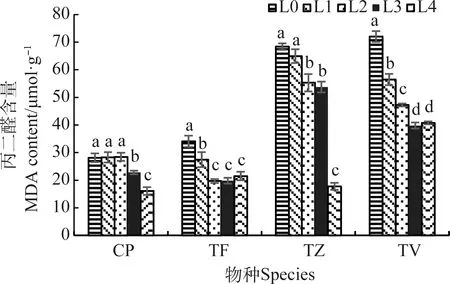
圖2 不同光強對丙二醛含量的影響
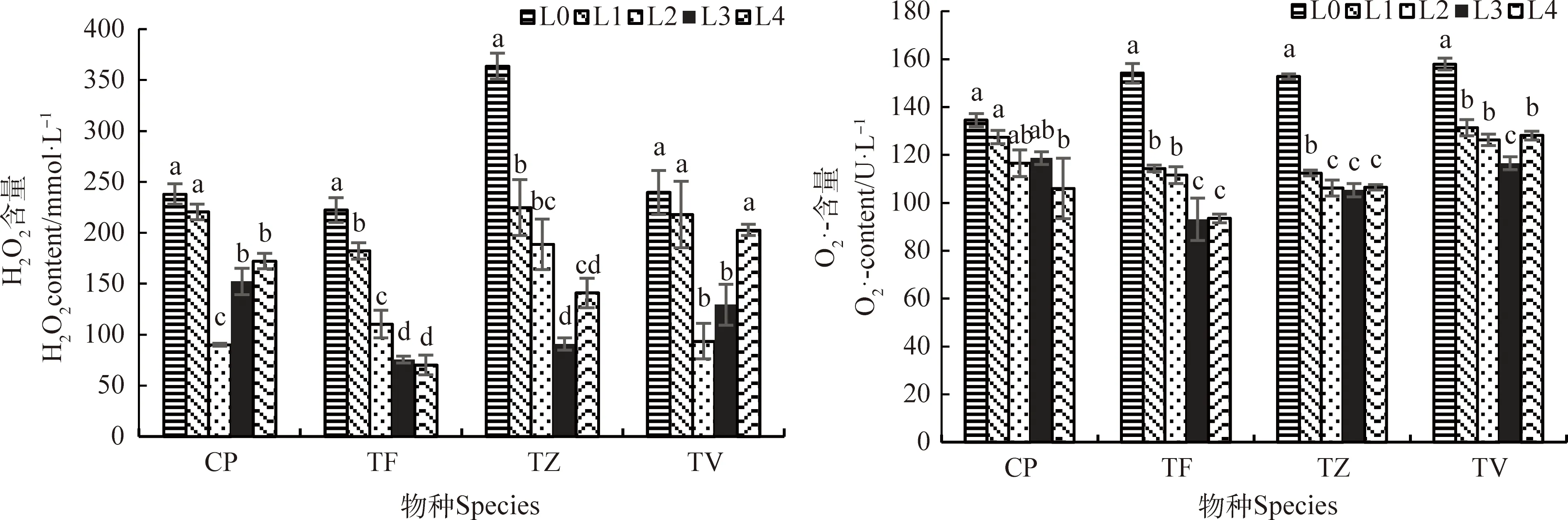
圖3 不同光強對H2O2和的含量影響

圖4 不同光強對葉片H2O2(DAB染色檢驗)和(NBT染色檢驗)的影響
2.4 光強對抗氧化酶活性的影響
隨光強的降低,CP和TV的SOD活性均呈“高-低-高”的變化趨勢(圖5),在L0條件下活性最高,且都在L3條件下降到最低值,與L0相比分別顯著降低了13.86%,17.93%(P<0.05);TF在L0和L1條件下的SOD活性顯著高于L2~L4條件(P<0.05),TZ在L0條件下的SOD活性顯著高于L1~L4條件(P<0.05)。
4種植物的CAT活性在不同的光強下呈不同的變化趨勢(圖5)。CP的CAT活性在各處理間無顯著差異,TF和TV的CAT活性隨光強的降低呈“高-低-高”的變化趨勢,其中,TF在L3條件下降到最低值,與L0相比,顯著將低了60.09% (P<0.05),TV在L2條件下降到最低值,與L0相比,顯著降低了35.49%;TZ的CAT活性與光強正相關,在L4條件下降到最低,與L0相比,顯著降低了40.63%。
4種植物的POD活性隨光強的降低顯著降低(P<0.05)(圖5),其中CP在L4條件下降到最低值,顯著降低了50.29%(P<0.05);TF,TZ和TV均在L3條件下降到最低值,與L0相比分別顯著降低了70.39%,56.80%,60.15%。
2.5 光強對葉肉細胞超微結構的影響
不同的光強對4種植物葉肉細胞的超微結構形態產生了不同程度的影響(圖6、圖7、圖8)。CP在L0條件下,細胞結構產生了嚴重的損傷,細胞膜解體并與細胞壁分離;葉綠體發生嚴重的形變,基粒片層和基質片層蜷曲,排列松散且片層間隙大,細胞器無規律的散布在細胞中,淀粉粒體積大,幾乎占據整個葉綠體。在L1~L4條件下,細胞質壁結構完好并結合緊密,葉綠體無溶解破碎現象,各片層結構排列緊密,呈紡錘形緊密貼合在細胞壁上,無堆疊擠壓的情況出現,淀粉粒體積也逐漸變小。不同的光環境對CP的葉綠體、淀粉粒、嗜鋨顆粒數量產生了顯著的影響。葉綠體個數隨著光強的減小而顯著增加(P<0.05),與L0相比,在L4條件下增加了39.57%;淀粉粒個數在L0條件下達到峰值,顯著高于其他光環境(P<0.05),是最低值L4條件的3.47倍;嗜鋨顆粒數量隨光強的降低呈先降低后增加的趨勢(圖8),在L3條件下數量最少;線粒體個數在不同的光環境下無顯著差異。
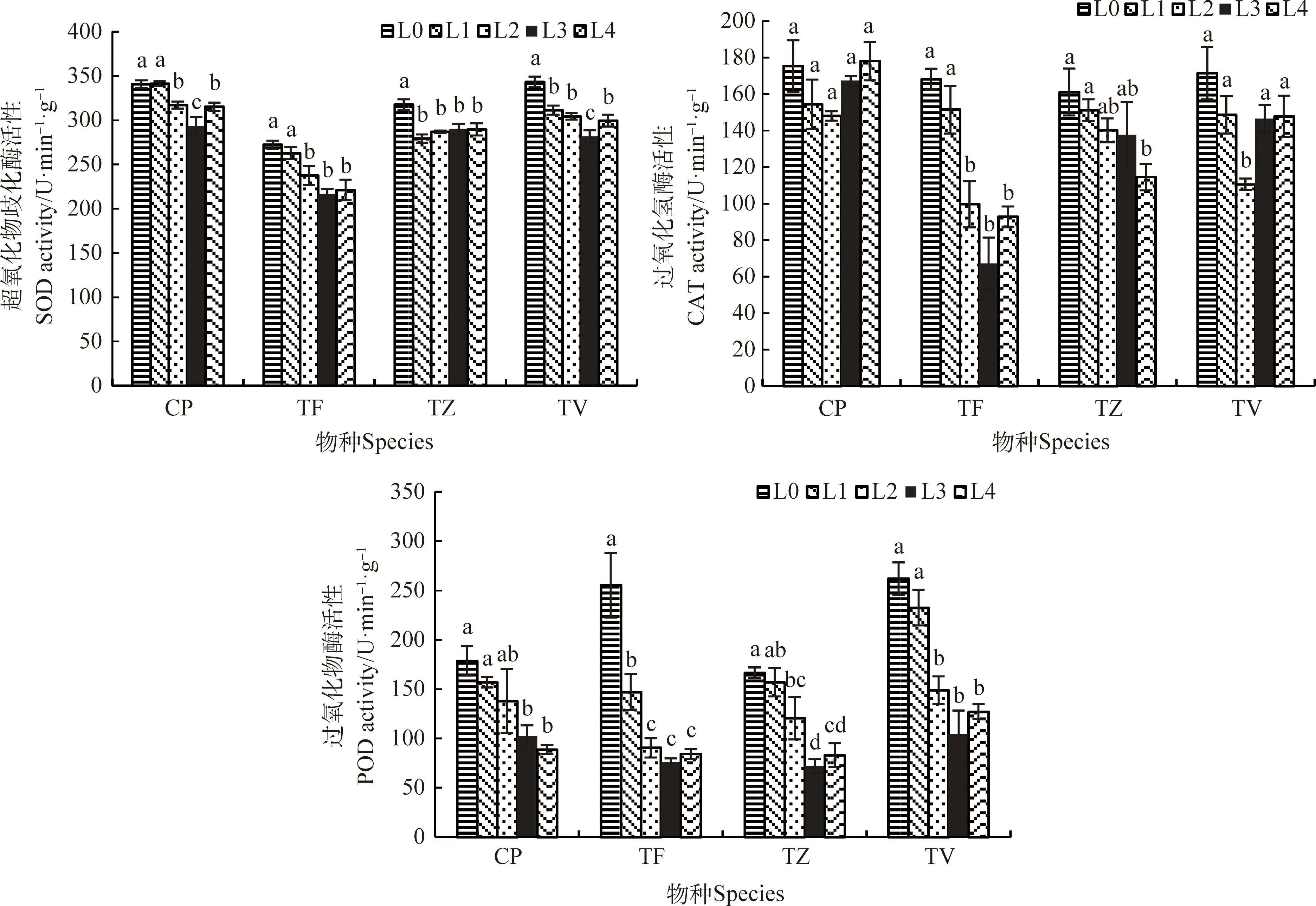
圖5 不同光強對抗氧化酶的影響
TF的細胞質壁結構在5種不同光強下結合緊密,界限清晰。在L0條件下,葉綠體部分解體,多個疊合分布在細胞壁邊緣,淀粉粒體積大,占據整個葉綠體結構的3/4左右;隨著光強的降低,淀粉粒體積逐漸變小,葉綠體也由寬圓形變為長紡錘形,單個獨立分布在細胞壁邊緣,但在L4條件下,部分葉綠體也發生了形變,雖然葉綠體在不同光環境下產生了不同的變化,但基質片層和基粒片層始終排列規則緊密。不同的光強對葉綠體、淀粉粒、線粒體和嗜鋨顆粒數量產生了顯著的影響(P<0.05)。L0條件下的葉綠體數量顯著低于其他光環境(P<0.05),在L3條件下達到峰值,而L0條件下的淀粉粒個數又顯著高于其他光照條件,其中淀粉粒個數在L3條件下降到最小值,嗜鋨顆粒隨光強的降低而顯著降低(P<0.05),在L3條件下數量最少,僅為L0條件下的34.78%,L1條件下的線粒體數量顯著高于其他光條件(P<0.05)。
TZ的細胞質壁結構在L0和L1條件下出現了嚴重的損壞,與細胞質界限不清,部分細胞膜解體,在L2,L3和L4條件下結構完好。L0和L1條件下的葉綠體結構破損,質膜溶解,部分基質片層和基粒片層結構出現解體現象,呈長圓形游離在細胞壁邊緣,淀粉粒體積大且出現空泡現象,在L2~L4條件下,葉綠體結構完好,片層結構緊密,緊貼細胞壁分布,淀粉粒體積也逐漸變小,在L4條件下消失。不同的光強對TZ的葉綠體、淀粉粒和嗜鋨顆粒數量產生了顯著的影響(P<0.05),與L0相比,在L4條件下葉綠體數量增加了30.73%,淀粉粒數量降低了79.15%,嗜鋨顆粒數量隨光強的降低而顯著降低,在L0條件下數量最多,在L2條件下最少,顯著降低了58.58%,線粒體數量在各處理下無顯著變化。
TV的細胞質壁結構在L0條件下發生了質壁分離的現象,但與細胞質界限清晰,在其他光環境下質壁結構完好。葉綠體在L0和L1條件下質膜解體,變為圓形,出現了堆疊現象,少量片層結構出現卷曲現象,但排列仍較為緊密,整個葉綠體被淀粉粒充斥。在L2~L4條件下,葉綠體結構完好,基質片層和基粒片層排列規則緊密,緊貼細胞壁分布,但仍有部分葉綠體發生形變現象,淀粉粒體積逐漸變小。不同的光強對TV的葉綠體、線粒體和嗜鋨顆粒數量產生了顯著的影響(P<0.05),葉綠體數量在L2條件下最多,在L4條件下最少。線粒體數量在L0條件下最少,顯著低于L1和L2條件(P<0.05),而其他處理間無顯著差異。嗜鋨顆粒數量隨光強的降低顯著降低(P<0.05),在L3條件下最少,與L0相比,顯著降低了48.42%。
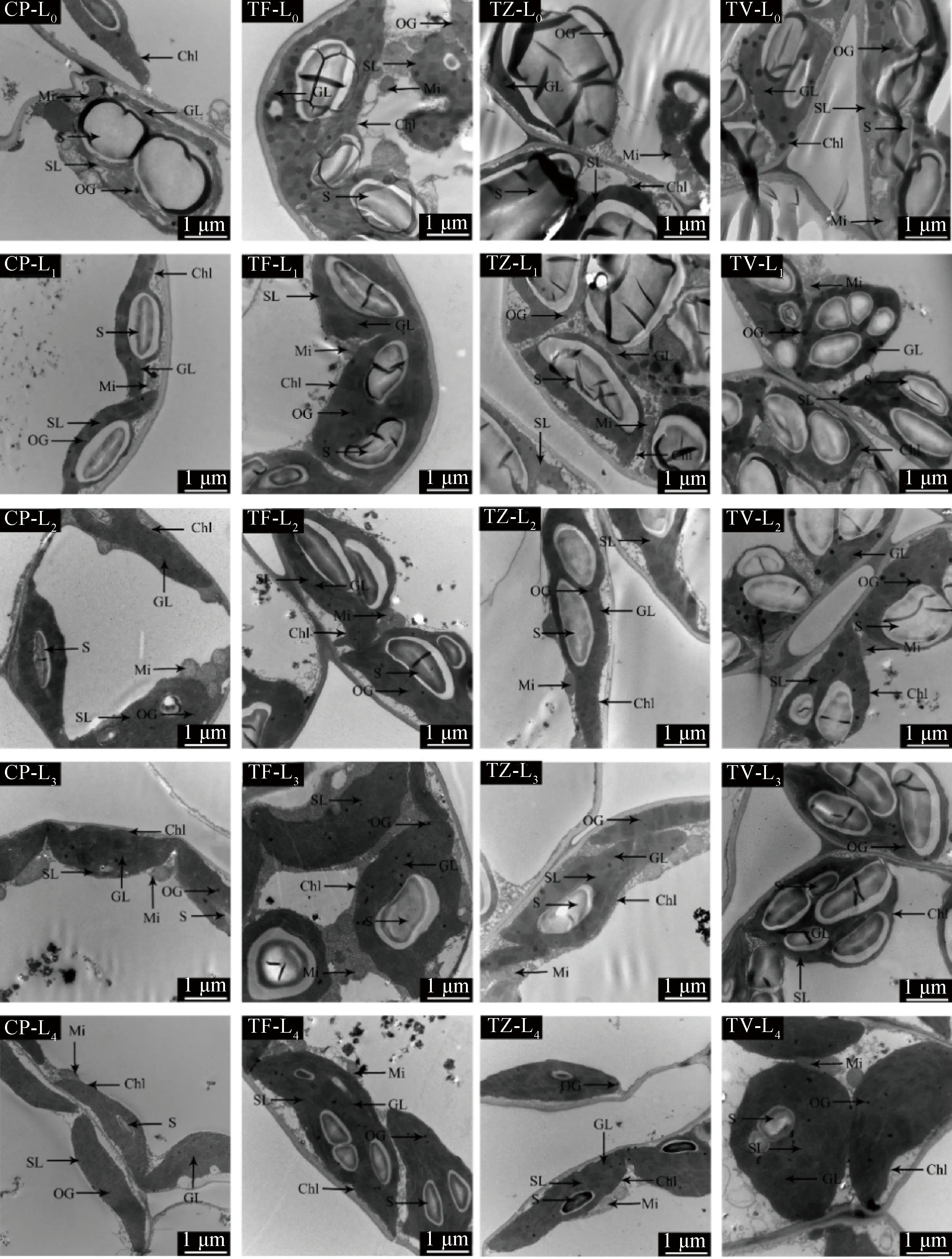
圖7 不同光強對葉綠體結構的影響(12000×)
2.6 物種和光強對不同指標的雙因素方差分析
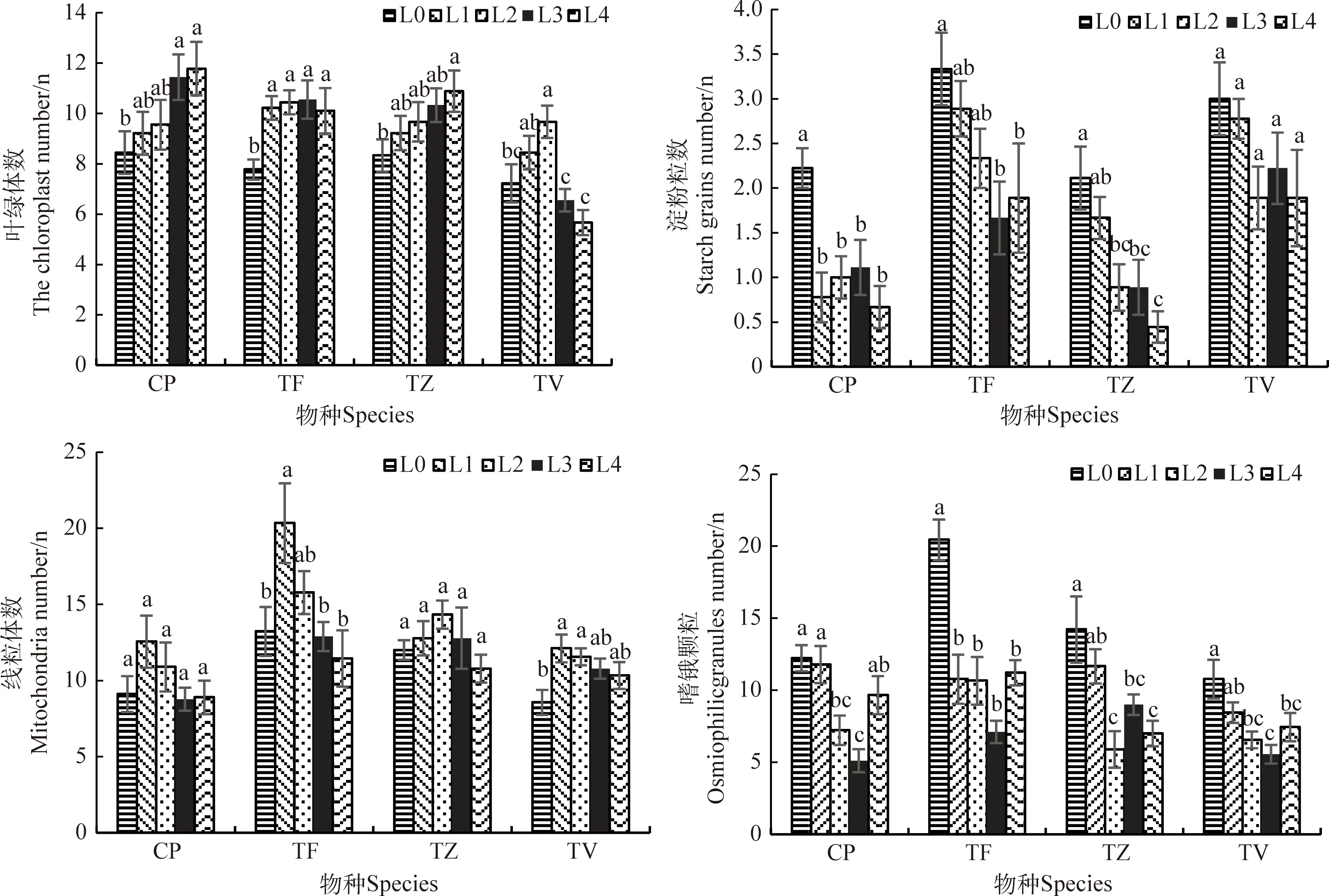
圖8 不同光強對葉肉細胞指標的影響
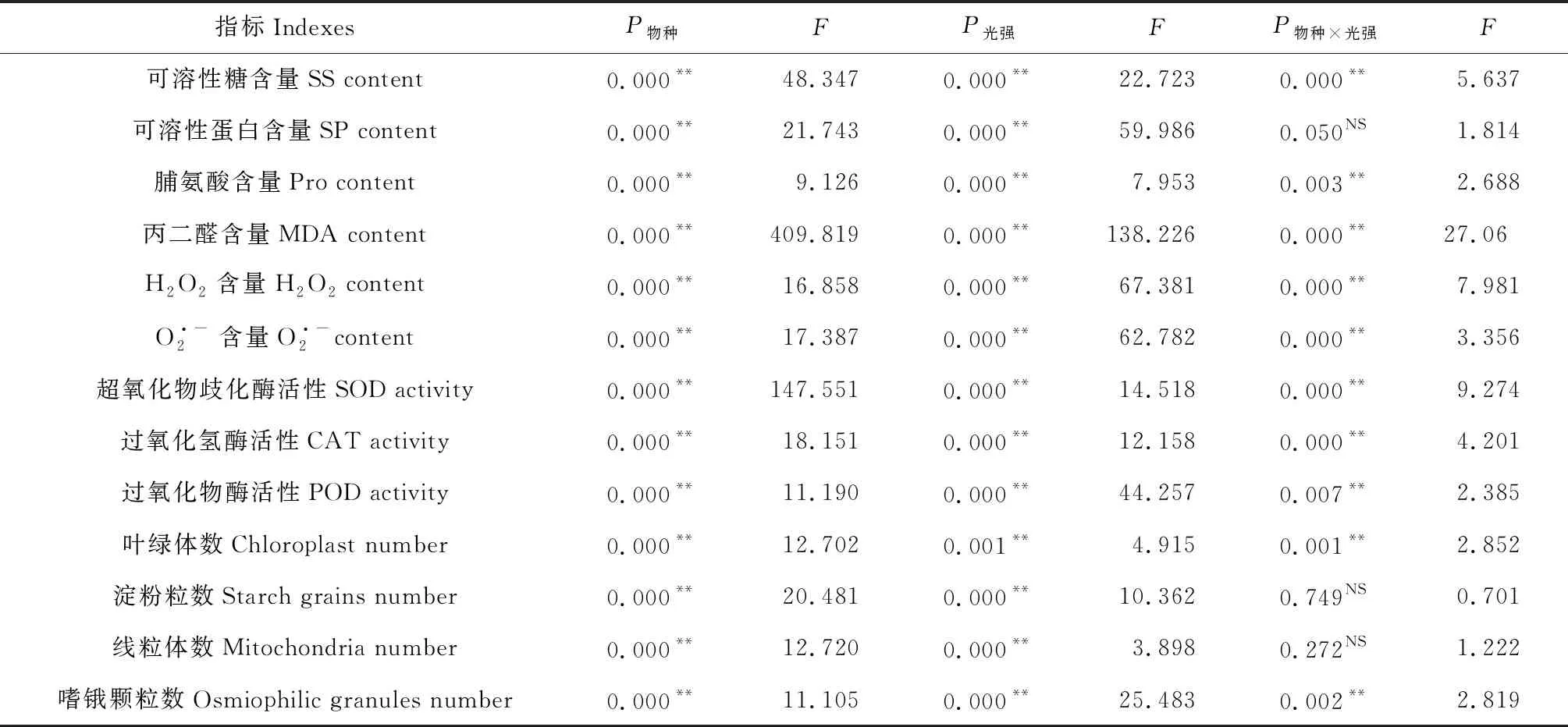
表2 物種和光強對不同指標的雙因素方差分析
2.7 4種植物的適光性綜合評價
通過隸屬函數法對4種鴨跖草科植物的適光能力進行了綜合評價(表3),結果表明適光性強弱的的順序為:吊竹梅>紫鴨跖草>‘花葉’水竹草>‘綠葉’水竹草。

表3 不同光強下4種植物生理指標的隸屬函數值
3 討論
滲透調節物質在植物抵抗非生物脅迫時發揮著舉足輕重的作用。當產生的活性氧超過抗氧化物清除能力時,細胞質膜會發生嚴重的膜脂過氧化反應,此時植物細胞膜的通透性就會變大,導致細胞質外滲,造成植物生理代謝紊亂[19],此時植物就會通過調整SS,SP,Pro等滲透調節物質的含量來減輕逆境脅迫引起的細胞失水,以維持細胞滲透壓平衡,從而降低逆境傷害的程度[20]。SS和SP既是重要的營養物質,又參與植物細胞的滲透調節過程[21],其中SS還可清除液泡和葉綠體中的ROS[22],可間接激活抗氧化酶系統。Pro被證明是一種有效的自由基清除劑,對穩定原生質膠體、細胞代謝過程、維持滲透調節平衡等方面起重要作用[23-24]。本研究發現‘花葉’水竹草和‘綠葉’水竹草的SS含量和SP含量均隨光強的降低而顯著降低,這表明這兩種植物在強光脅迫下均能通過增加SS含量和SP含量,以清除液泡和葉綠體中的ROS,并通過碳代謝途徑和氮代謝途徑協同作用進行滲透調節,維持細胞膨壓;而紫鴨跖草和吊竹梅的SS含量隨著光強的降低呈先升高后降低的趨勢,SP含量則隨光強的降低而顯著降低,這表明這2種植物在強光脅迫下氮代謝途徑對滲透調節的貢獻比碳代謝途徑大,而在75%~25%自然光強中,碳代謝途徑對滲透調節的貢獻大于氮代謝途徑。4種植物的SS含量和SP含量均在L4條件下降到最低,這可能是由于光強不足,葉片的光合作用急劇下降,SS和SP合成受阻,最終會導致植物的營養物質短缺,對生長造成不利影響。‘花葉’水竹草和‘綠葉’水竹草的Pro含量在各處理間無顯著差異,這表明這2種植物的Pro含量穩定,始終保持相對較高的含量,清除植物細胞內的自由基,并抵消光強因素對細胞滲透調節的影響,而紫鴨跖草和吊竹梅的Pro含量隨光強的降低呈先降后增的趨勢,這表明Pro在極端弱光中仍能很好的發揮滲透調節的作用。

4 結論
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
