苜蓿抗寒性與根系抗氧化酶活性相關性分析
2021-04-08 10:55:44張玉霞叢百明王顯國張慶昕杜曉艷田永雷
草地學報 2021年2期
張玉霞, 叢百明, 王顯國, 張慶昕, 杜曉艷, 田永雷
(1.內蒙古民族大學, 內蒙古 通遼 028041;2.內蒙古通遼市農牧業局,內蒙古 通遼 028000;3. 中國農業大學草業科學與技術學院, 北京 100083;4.內蒙古自治區農牧科學院,內蒙古 呼和浩特 010013)
苜蓿作為優質的多年生豆科牧草(Leguminosaesp.)[1],在畜牧業生產中發揮著重要作用[2]。越冬問題雖然是長期困擾苜蓿生產的老問題,但由于近幾年我國北方苜蓿種植面積不斷擴大,苜蓿產業發展迅猛,越冬問題已成為制約我國北方苜蓿草地成功建植和草地可持續利用的關鍵問題。由于科爾沁沙地冬季積雪少,土壤溫度變化劇烈,凍害頻發,嚴重制約了該地區苜蓿產業的發展[3]。因此,進一步探究苜蓿的越冬機制,以增加苜蓿防御凍害的能力,提高其越冬率,重新認識苜蓿抗寒性問題是必要的。
國內外關于苜蓿抗寒方面的研究表明,苜蓿越冬器官的超氧化物歧化酶(Superoxide dismutase,SOD)、過氧化物酶(Peroxidase,POD)、過氧化氫酶(Catalase,CAT)是酶促防御系統的重要保護酶類,對于維護細胞膜系統的穩定性具有重要作用[4-6],其活性的變化可作為植物的耐寒指標[7-9],但具體什么時期或在怎樣溫度條件下進行鑒定尚不明確。目前衡量苜蓿抗寒性主要指標有大田越冬率調查和室內模擬半致死溫度(LT50),LT50采用模擬低溫處理通過測定相對電導率或根系活力擬合Logistic方程獲得,能夠避開生態因子對苜蓿抗寒性的影響[10]。為此,本試驗于越冬前期挖取科爾沁沙地生境下種植的黃花苜蓿(MedicagofalcataL.)和4個紫花苜蓿(Medicagosativa)品種的越冬器官,在程序式溫度處理箱模擬低溫處理,測定不同溫度處理下抗氧化酶活性和相對電導率的變化,利用相對電導率擬合Logistic方程得出LT50,通過LT50與抗氧化酶活性變化的相關性分析,分析苜蓿的抗寒性差異及其與抗氧化酶的相關性,為苜蓿抗寒品種的篩選及抗氧化機理研究提供理論依據。
1 材料與方法
1.1 供試材料
苜蓿材料來源見表1。

表1 苜蓿材料名稱及來源
1.2 試驗地概況
試驗地位于內蒙古民族大學科技園區,地理位置為43°30′ N,122°27 E,年平均氣溫5.5℃,≥10℃年積溫為3 000~3 200℃,無霜期140~150 d,年平均降水量約375 mm,年平均風速3.0~4.4 m·s-1,土壤為沙壤土,pH值為8.0,土壤有機質含量0.65%,堿解氮36.37 mg·kg-1,速效磷含量3.81 mg·kg-1,速效鉀含量78.51 mg·kg-1,全氮含量0.037%。
1.3 試驗設計
采用隨機區組試驗設計,在科爾沁沙地的沙化草地生境下,于2019年7月15日種植黃花苜蓿和‘騎士T’、‘巨能3010’、‘草原3號’、‘北極熊’紫花苜蓿,播種量15 kg·hm-2,條播行距15 cm,小區面積(2 m×5 m),3次重復,四周設保護行。灌溉方式為指針式噴灌,肥料在播種時一次性施入,氮磷鉀施肥量分別為45 kg·hm-2(純氮)、200 kg·hm-2(P2O5)、200 kg·hm-2(K2O),供試肥料分別為尿素(N 46.3%)、重過磷酸鈣(P2O544.6%)、鉀肥為氯化鉀(K2O 60%)。2019年11月15日(封凍前期)每個小區隨機挖取50株長勢均勻的苜蓿越冬器官,4℃冰箱中儲存備用。每個處理分別取20株于程序式低溫處理箱采用-10,-15,-20,-25和-30℃模擬低溫處理,以0℃為起點,4℃·h-1的速率降溫,到達設定溫度后保持6 h,然后再按照4℃·h-1速率升溫至4℃,在冰箱4℃下保存12 h,以低溫(4℃)貯藏為對照(CK),測定抗氧化酶活性和相對電導率,并依據測定其相對電導率擬合Logistic方程,計算半致死溫度。
1.4 測定指標及方法
采用DDS-302電導率儀測定電導率并計算相對電導率[11];采用NBT光還原法測定SOD活性[11];采用愈創木酚法測定POD活性[12];采用紫外吸收法測定CAT活性[11]。
1.5 數據分析
試驗數據用Microsoft Excel軟件處理、做圖和制作表格,SPSS 17.0軟件進行方差顯著性分析及擬合Logistic方程。
2 結果與分析
2.1 低溫脅迫下苜蓿相對電導率的變化及LT50
如表2所示,隨著脅迫溫度的降低,苜蓿根頸的相對電導率呈增加趨勢,其中‘騎士T’、‘巨能3010’、‘草原3號’和‘北極熊’等紫花苜蓿品種根頸的相對電導率則在-15℃低溫處理開始顯著增加(P<0.05);黃花苜蓿的相對電導率則在-20℃低溫處理開始顯著增加(P<0.05)。在CK和-10℃低溫處理下不同苜蓿之間的相對電導率無顯著差異,-15℃低溫處理下黃花苜蓿根頸相對電導率顯著低于‘巨能3010’和‘北極熊’(P<0.05),-20℃處理下黃花苜蓿和‘北極熊’相對電導率顯著低于‘騎士T’和‘巨能3010’(P<0.05),-25℃和-30℃低溫處理下黃花苜蓿根頸的相對電導率則分別顯著低于‘騎士T’和‘巨能3010’(P<0.05)。根據苜蓿根頸相對電導率結合Logistic回歸分析,計算半數致死溫度,黃花苜蓿的抗寒性強于紫花苜蓿,4個紫花苜蓿品種的抗寒性為‘北極熊’>‘草原3號’>‘巨能3010’>‘騎士T’。
2.2 低溫脅迫下不同苜蓿根頸SOD活性的變化
由表3可知,在不同低溫處理下,‘騎士T’、‘巨能3010’、‘草原3號’苜蓿根頸的SOD活性呈先增加后降低的變化趨勢,且均在-25℃低溫處理下苜蓿根頸的SOD活性最強,與—30℃處理差異不顯著。‘北極熊’和黃花苜蓿根頸SOD活性則隨著處理溫度的降低呈不斷增加趨勢,其中‘騎士T’和黃花苜蓿則在-20℃處理下開始顯著增加,‘巨能3010’、‘草原3號’和‘北極熊’在-25℃處理開始顯著增加。-20℃低溫處理下黃花苜蓿根頸SOD活性顯著高于‘巨能3010’(P<0.05),-25℃處理下黃花苜蓿根頸SOD活性顯著高于‘騎士T’(P<0.05)。—30℃處理下黃花苜蓿的SOD活性顯著高于‘騎士T’、‘巨能3010’、‘草原3號’。因此-25℃和-30℃低溫處理下苜蓿根頸的SOD酶活性可以作為鑒定苜蓿抗寒性的生理指標。

表2 低溫脅迫下苜蓿根頸相對電導率(%)的變化及LT50值(℃)

表3 低溫脅迫下苜蓿根頸SOD活性的變化
2.3 低溫脅迫下不同苜蓿根頸POD的變化
由表4可知,黃花苜蓿和4個紫花苜蓿品種在低溫脅迫下根頸POD活性均呈先升高再降低的趨勢。‘騎士T’苜蓿根頸的POD活性在-15℃處理下最高,顯著高于對照和-30℃處理(P<0.05),‘巨能3010’苜蓿根頸的POD活性在-20℃處理下最高,顯著高于其他溫度處理(P<0.05),‘草原3號’、‘北極熊’和黃花苜蓿根頸的POD活性在-25℃處理下最高,顯著高于其他溫度處理(P<0.05)。-20℃,-25℃,-30℃低溫處理下黃花苜蓿根頸的POD活性顯著高于4個紫花苜蓿品種(P<0.05)。由此說明,抗寒性強的苜蓿材料POD活性增強溫度較低,且活性較強。
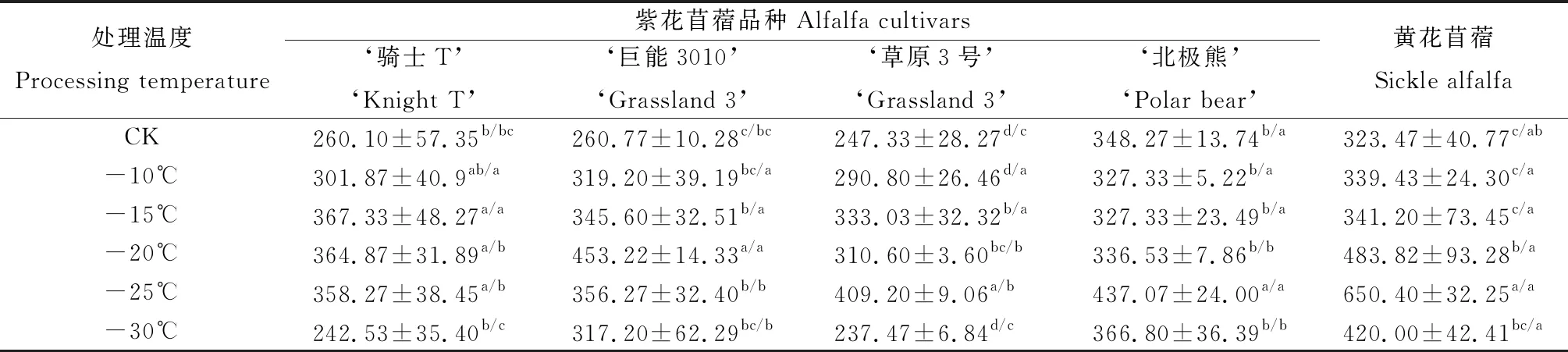
表4 低溫脅迫下苜蓿根頸POD活性的變化
2.4 低溫脅迫下不同苜蓿根頸CAT的變化
由表5可知,黃花苜蓿和4個紫花苜蓿品種根頸CAT活性隨脅迫溫度的降低呈先降低再升高的變化趨勢。-10℃低溫處理下‘草原3號’根頸CAT活性顯著低于其他溫度處理(P<0.05),黃花苜蓿和其他3個紫花苜蓿品種在-20℃低溫處理下根頸CAT活性最顯著低于-30℃低溫處理(P<0.05)。-15℃和-20℃低溫處理下‘草原3號’根頸CAT活性顯著高于黃花苜蓿和其他3個紫花苜蓿品種(P<0.05),-25℃低溫處理下黃花苜蓿根頸的CAT活性顯著高于‘騎士T’、‘巨能3010’、‘北極熊’,-30℃低溫處理下黃花苜蓿根頸CAT活性顯著高于4個紫花苜蓿品種(P<0.05)。說明‘草原3號’和黃花苜蓿通過增強CAT活性提高苜蓿的抗寒能力。
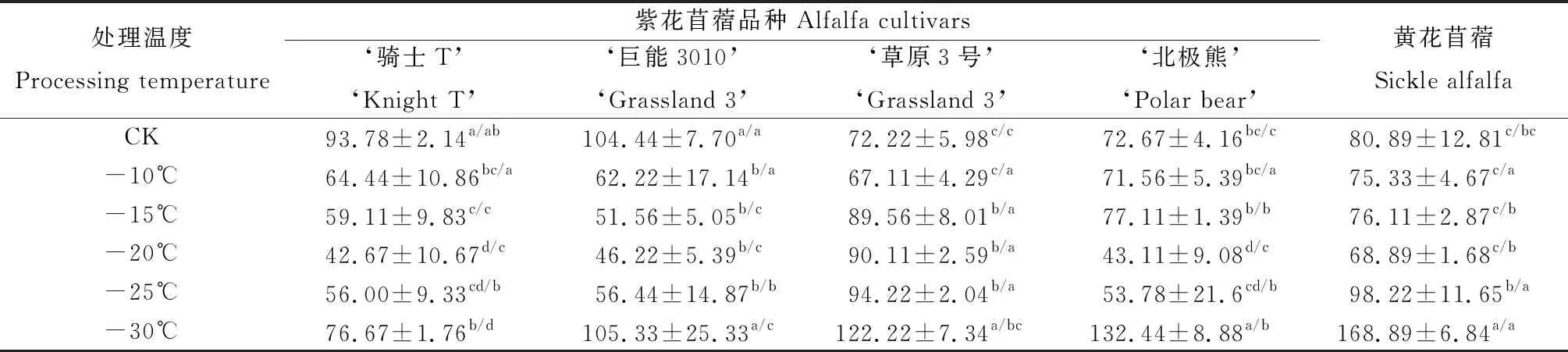
表5 低溫脅迫下苜蓿根頸CAT活性的變化
2.5 低溫脅迫下苜蓿根頸抗氧化酶活性與LT50的相關性分析
由表6可知,不同低溫脅迫下苜蓿的抗氧化酶活性均與LT50呈負相關,—30℃低溫處理下苜蓿根頸SOD和CAT活性與其LT50的相關性達到顯著(P<0.05)和極顯著水平(P<0.01),說明—30℃低溫脅迫下SOD活性和CAT活性可以作為判斷苜蓿抗寒性強弱的敏感指標。

表6 半致死溫度與苜蓿根頸中抗氧化酶活性的相關性
3 討論
關于苜蓿抗寒性的研究主要采用田間自然低溫脅迫處理的方法[7],通過測定翌年的越冬率衡量苜蓿抗寒性的強弱,抗寒生理指標則采用越冬期不同低溫處理時期取樣測定,這種方法容易受外界氣候和土壤環境影響,冬季取樣困難。近些年開始采用程序式試驗箱人工模擬低溫處理方法[13],利用半致死溫度衡量苜蓿的抗寒性,這種方法能夠避開其他環境因素的影響,取樣測定抗寒生理指標簡便易行。本研究則通過大田培養苜蓿材料,采用人工模擬低溫處理方法,獲得半致死溫度判定苜蓿抗寒性差異。結果表明,黃花苜蓿的抗寒性強于紫花苜蓿,4個紫花苜蓿品種的抗寒性為‘北極熊’>‘草原3號’>‘巨能3010’>‘騎士T’。
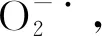
POD在保護酶系統中主要是降解過氧化物的作用[14],關于POD活性在低溫脅迫下的變化規律,申曉慧研究田間自然低溫脅迫下POD活性的變化,表明隨著氣溫的下降而上升,進入深冬又下降,翌年春季氣溫回升后,又有所回升[7];崔國文研究表明,POD活性都表現為隨著秋末溫度的下降而增加,隨著寒冷的加劇而迅速下降,再隨著翌年溫度的回升而緩慢增加,但POD活性與品種抗寒力不呈正相關關系[15]。馮昌軍研究表明,隨著低溫脅迫時間的延長,POD活性呈先增加后降低的變化趨勢,抗寒性強的苜蓿品種維持較高的POD酶活性[16]。本研究采用人工模擬低溫處理方法研究苜蓿根頸的POD變化規律,結果表明,隨著脅迫溫度的降低,苜蓿根頸的POD活性呈先增加后降低的變化趨勢,與申曉慧[7]、崔國文[15]、馮昌軍[16]的研究結果一致,但與崔國文研究不同的是,苜蓿根頸的POD活性與苜蓿的抗寒性呈正相關,這與南麗麗[17]、申曉慧[7]、朱愛民[18]的研究結果一致,通過與苜蓿抗寒性的相關性分析表明,苜蓿根頸的POD活性與苜蓿抗寒性的相關性不顯著,因此POD活性不能作為鑒定苜蓿抗寒性的生理指標。
CAT具有將低溫脅迫下產生的H2O2分解成水的作用[19]。關于低溫脅迫下苜蓿根頸CAT活性的變化規律研究較少,楊秀娟采用田間自然低溫脅迫處理研究,表明2個紫花苜蓿品種根頸中CAT活性均隨著溫度的降低而呈現先上升后下降的趨勢,在初冬時達到最大值,進入深冬,則有不同程度的波動,且活性下降明顯[20]。本研究采用模擬低溫脅迫處理,研究苜蓿根頸CAT活性的變化,與其結果不同,隨著冷凍脅迫溫度的降低,苜蓿的CAT活性呈增加的變化趨勢,在—30℃低溫脅迫下黃花苜蓿和4個紫花苜蓿品種根頸的CAT活性最強,相關性分析表明,在-10℃和—30℃低溫處理下苜蓿根頸的CAT活性與苜蓿的抗寒性呈顯著正相關,說明-10℃和—30℃低溫處理下苜蓿根頸的CAT活性都可以作為苜蓿抗寒性的鑒定指標。
4 結論
科爾沁沙化草地種植的苜蓿抗寒性為黃花苜蓿強于紫花苜蓿,4個紫花苜蓿品種的抗寒性為‘北極熊’>‘草原3號’>‘巨能3010’>‘騎士T’。LT50與不同低溫脅迫下抗氧化酶活性相關性分析表明,—30℃低溫處理下SOD活性和CAT活性以及-10℃低溫處理下的CAT活性可以作為苜蓿抗寒鑒定的敏感指標。
