ADAMTS9基因啟動子甲基化在直腸癌中的表達與臨床病理特征的相關性
2021-03-22 06:40:36高永濤
實用癌癥雜志 2021年3期
劉 濤 白 浪 高永濤
直腸癌是消化系統常見的惡性腫瘤,在中國的發病率也呈逐年上升趨勢,嚴重威脅人類健康[1-3]。多項研究證實直腸癌的發生發展與癌基因、抑癌基因突變有關[4]。抑癌基因甲基化是腫瘤發生發展的常見的表觀遺傳學變化,有研究證實抑癌基因啟動子區CpG島甲基化與直腸癌的發生發展有關[5]。ADAMTS(a disintegrin and metalloproteinase with thrombospondin motifs,ADAMTS)是一種新型金屬蛋白酶家族,參與多種生理學和病理學過程如血管生成、癌癥發生等[6]。ADAMTS9是ADAMTS家族成員之一,作為抑癌基因抑制多種腫瘤發生發展[7]。本研究旨在探討ADAMTS9基因甲基化在直腸癌發生發展中的作用。
1 材料與方法
1.1 材料
1.1.1 主要試劑 TRIzol(美國Invitrogen公司)、DNA提取試劑盒(德國Qiagen公司)、Taq酶(日本Takara公司)、CpGenome DNA修飾試劑盒(Chemicon公司)。
1.1.2 標本 在取得直腸癌患者或家屬知情同意后,收集2016年1月至2018年12月間于延安大學附屬醫院普通外科醫院行直腸癌根治手術切除的83例直腸癌組織及其相應的癌旁組織標本。所有患者直腸標本均經術后病理證實直腸腺癌,術前均未行新輔助放化療,83例直腸癌手術中,57例行腹腔鏡直腸癌根治術,26例行開腹直腸癌根治術。83例患者中,男性52例,女性31例,平均年齡(53.8±10.2)歲。
1.2 方法
1.2.1 提取直腸癌組織、癌旁組織中RNA、DNA 在液氮中將直腸癌和癌旁組織研磨至粉末狀,加入1 ml的TRIzol試劑后依次提取細胞總RNA和基因組DNA。
1.2.2 ADAMTS9 mRNA表達水平檢測 利用Real Time PCR檢測ADAMTS9 mRNA表達水平,分別取上述直腸癌、癌旁組織總RNA 2 μg進行逆轉錄,采用Takara公司SYBR Premix Ex TaqTMⅡ試劑盒,以β-actin為內參,測定ADAMTS9基因的mRNA水平。ADAMTS9上游引物:5'-GGATCGTGGCTACTGACCGCAA-3',下游引物:5'-GACGTTGACGCCGATTGCACTAA-3'。反應體系包括:SYBR Green I Master Mix 12 μl、10 μmol/l引物各1 μl、cDNA 2 μl,加去離子水補足體積至25 μl。反應條件:52 ℃ 1 min,95 ℃預變性3 min,40個循環,目的基因相對于內參基因的mRNA水平以2-ΔΔCt值表示。
1.2.3 ADAMTS9基因啟動子甲基化分析 分別取上述直腸癌、癌旁組織總DNA,采用CpGenome修飾試劑盒進行亞硫酸氫鈉修飾,利用巢式-甲基化特異性PCR(nest-methylation-specific PCR,MSP)進行ADAMTS9基因啟動子區甲基化水平的檢測。ADAMTS9上游引物:5'-AACGTTGACGTGACCGGTACGACT-3',下游引物:5'GGCATTGACTCAAGTCG-3',USP上游引物:5'-TTGCAACGGCATGACC-3',下游引物:5'-GGCATGACTTACGGTTACGCATC-3'。^MSP反應體系如下:總RNA 1 μl,PCR buffer 2.5 μl,引物各0.5 μl,加至總體積25 μl,反應條件:94 ℃預變性5 min,94 ℃ 30 s,55 ℃ 60 s,共 35個循環,72 ℃延伸10 min。取10 μl的PCR擴增產物進行3%瓊脂糖凝膠電泳,ChemilmagerTM5500凝膠成像處理系統觀察電泳結果。
1.3 統計方法
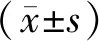
2 結果
2.1 直腸癌、癌旁組織中ADAMTS9 mRNA表達水平
通過Real Time PCR 檢測ADAMTS9基因在83例直腸癌組織和配對癌旁組織中的mRNA相對表達水平,結果顯示直腸癌組織中ADAMTS9 mRNA相對表達水平為(23.6±8.59),低于配對癌旁組織的(35.2±9.18),差異有統計學意義(χ2=1161,P<0.0001),表明ADAMTS9基因可能在mRNA轉錄水平參與直腸癌的發生發展過程。見圖1。

圖1 直腸癌、癌旁組織中ADAMTS9 mRNA表達水平
2.2 直腸癌、癌旁組織中ADAMTS9基因啟動子甲基化狀態
應用MSP方法分析83對直腸癌組織和癌旁組織的ADAMTS9基因啟動子區CpG島的甲基化狀態(圖2),結果顯示83例直腸癌組織中ADAMTS9基因啟動子區CpG島甲基化17例,甲基化發生率20.5%,83例癌旁組織中ADAMTS9基因啟動子區CpG島甲基化發生率7.2%(6/83),直腸癌組織與癌旁組織的ADAMTS9基因啟動子區CpG島的甲基化率有顯著性差異(χ2=37.19,P<0.0001)。Logistic回歸分析顯示ADAMTS9基因啟動子區CpG島的異常甲基化可增加直腸癌發病風險(OR=10.254,95%CI=3.395~25.631)。

圖2 ADAMTS9基因啟動子區在直腸癌和癌旁組織中的甲基化電泳圖譜
2.3 ADAMTS9基因啟動子甲基化與直腸癌臨床病理特征之間的相關性
通過分析直腸癌組織中ADAMTS9啟動子區甲基化頻率與直腸癌臨床病理特征的相關性,結果表明直腸癌組織中ADAMTS9啟動子區甲基化與腫瘤TNM分期、分化程度相關。T1、T2,高分化,Ⅰ、Ⅱ期直腸癌患者中ADAMTS9啟動子區甲基化頻率更低,而ADAMTS9啟動子區甲基化與直腸癌患者年齡、性別、腫瘤大小無明顯相關性,表明ADAMTS9啟動子區甲基化與直腸癌發生發展有關。見表1。

表1 ADAMTS9啟動子區甲基化與直腸癌臨床病理特征之間的相關性(例,%)
3 討論
直腸癌是我國常見的消化道惡性腫瘤之一,近年來其發病率和死亡率均呈逐年上升趨勢,因此,研究直腸癌的發病機制具有很高的價值[8-9]。隨著對直腸癌發病機制的研究不斷深入,大量研究證實直腸癌的發生與遺傳和環境長期作用密切相關,癌基因和抑癌基因的失衡是直腸癌發病的重要機制[10-11]。
越來越多的證據表明表觀遺傳異常也在直腸癌的發病中發揮作用。DNA甲基化(DNA methylation)是表觀遺傳學的重要現象,與基因組印記(genomic imprinting),基因沉默(gene silencing)等表觀遺傳現象參與腫瘤的發生發展[12-15]。基因組中DNA的甲基化模式是通過DNA甲基轉移酶實現哺乳動物基因表達調控的重要形式,DNA甲基化異常引起抑癌基因失活和癌基因激活是直腸癌發生的重要機制之一,DNA異常甲基化的逆轉可能有助于直腸癌的診斷和治療[16-17]。人類 ADAMTS 家族包含 19 個分泌型的多域蛋白水解酶,ADAMTS 家族成員作為腫瘤抑制基因(tumor suppressor genes,TSGs)發揮作用,有研究報道 ADAMTS家族成員的表觀遺傳學修飾參與直腸癌發病機制[18]。有研究表明,ADAMTS9在食管癌組織中表達降低,且與ADAMTS9基因的啟動子甲基化有關[19-20]。但研究ADAMTS9基因在直腸癌中的作用較少。本研究發現,直腸癌組織中ADAMTS9 mRNA表達水平低于配對癌旁組織(χ2=1161,P<0.0001),表明ADAMTS9基因可能在mRNA轉錄水平參與直腸癌的發生發展過程。通過應用MSP方法,進一步分析ADAMTS9基因啟動子區CpG島的甲基化狀態,結果直腸癌組織中ADAMTS9基因啟動子區CpG島甲基化發生率為20.5%,高于癌旁組織甲基化發生率7.2%(χ2=37.19,P<0.0001)。Logistic回歸分析顯示ADAMTS9基因啟動子區CpG島的異常甲基化可增加直腸癌發病風險。分析直腸癌組織中ADAMTS9啟動子區甲基化頻率與直腸癌臨床病理特征的相關性,結果表明直腸癌組織中ADAMTS9啟動子區甲基化與腫瘤TNM分期、分化程度相關,進一步證實抑癌基因ADAMTS9啟動子區甲基化不但參與了直腸癌的發生,而且與直腸癌的進展和轉移有關。上述研究結果提示ADAMTS9啟動子區甲基化可為直腸癌的早期診斷及預后判斷提供幫助。然而,關于ADAMTS9啟動子區甲基化如何進一步影響直腸癌發生發展的機制仍有待于進一步研究。
直腸癌的發生、發展是多個基因、多種因素綜合作用的結果,隨著對DNA甲基化和表觀遺傳學研究的深入,我們終將揭開直腸癌的發病機制。我們的研究初步證實了直腸癌組織中抑癌基因ADAMTS9啟動子區異常甲基化與腫瘤發生發展密切相關,為后續直腸癌的表觀遺傳學研究提供了依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
火花(2019年12期)2019-12-26 01:00:28
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
汽車工程學報(2017年2期)2017-07-05 08:13:02
學苑創造·A版(2015年11期)2016-01-14 09:03:27
