黏性魚卵的黏性機制及人工孵化技術研究進展
2021-03-13 06:48:04高鳳祥胡發文菅玉霞李莉王雪王曉龍潘雷郭文
大連海洋大學學報 2021年1期
高鳳祥,胡發文,菅玉霞,李莉,王雪,王曉龍,潘雷,郭文
(山東省海洋生物研究院 青島市海洋生物種質資源挖掘與利用工程實驗室,青島淺海底棲漁業增殖重點實驗室,山東 青島 266104)
魚類的繁殖策略是為了適應其生活的水環境棲息地。魚卵是魚類的雌性生殖細胞,其產出后黏性程度因種類的不同有較大差異,Rizzo等[1]通過對魚卵的宏觀評估,將魚卵按黏性程度分為黏性卵、弱黏性卵和非黏性卵3類。其中,緊密黏附在一起形成黏性卵塊的魚卵為黏性卵,彼此黏附但在輕微攪動下可自由移動的卵為弱黏性卵,而完全自由游離的卵為非黏性卵。產黏性卵的代表魚類有大瀧六線魚Hexagrammosotakii、松江鱸Trachidermusfasciatus、絨杜父魚Hemitripterusvillosus等,這類魚卵卵膜表層遇水后表現出極高黏性,呈團塊狀附著于水草、海藻或巖礁等產卵基質上,一般具有黏性強、孵化時間長等特點;產弱黏性卵的代表種類有西伯利亞鱘Acipenserbaerii等;還有一些魚類所產的卵無黏性,卵粒分離,能自由浮動[2],代表性種類有大菱鲆Scophthalmusmaximus等。
魚卵有無黏性與魚的生活史和繁殖策略有重要關系。在自然條件下,遷徙魚種的卵通常無黏性,會自由地漂向下游。定居魚種的卵通常具有黏性,會附著在產卵地附近不同的底層物質上[3]。因此,卵的黏性特征可能有助于理解不同種類硬骨魚間的系統發育關系[4]。黏性魚卵遇水后,表面黏性物質即被激活,魚卵或黏附在基質上或互相黏連成團,不同魚類卵的黏性成因和黏度各有不同。經長期大量的試驗和生產實踐,科研人員研發了多種黏性魚卵人工授精和孵化方法。本研究中,綜述了黏性魚卵表面黏性物質的結構成分及黏連機制,綜合對比多種黏性卵人工授精及孵化方法,分析了這些方法的原理及優缺點,并對今后的研究方向進行了思考且提出相應建議,以期為產黏性卵魚類的人工繁育提供參考。
1 魚卵黏性的生物學功能
魚卵的黏性是為適應生活環境而發展出的繁殖策略。許多硬骨魚類會產下黏性卵,卵子遇水后迅速出現的黏性是防止卵從最佳棲息地漂移到次等棲息地的重要機制[5]。魚卵的黏性有助于其附著在基質上,或與其他卵相互黏附,以防止魚卵被水流、海浪沖散或被捕食者捕食。魚卵沉降附著在固定的領地內有助于親魚護卵,提高存活率[6]。在上層和底棲魚類中,魚卵的附著方式和黏附程度各不相同,這種繁殖策略的變化反映了不同魚類對環境條件的適應性。
鱘的魚卵在與水接觸后的幾分鐘內便會產生黏性,具體時間因種類而異,如俄羅斯鱘Acipensergueldenstaedti為8~19 min,歐洲鰉Husohuso為4~6 min,閃光鱘Acipenserstellatus為5~12 min[7]。鱘類中有5種產出弱黏性卵,有22種產出黏性卵,其中有3種卵黏性極強。大多數種類的鱘卵是沉黏性卵,產在水底的泥灘、沙地或巖石上的沙礫、鵝卵石這類基質上[8-9]。所有種類的鱘均會產出具有黏性的卵,使得魚卵可以附著在基質上。Vorob’eva等[10]推測,鱘卵黏性的作用很可能是將卵固著在潔凈的基質上以便助其受精,進而提高自然條件下鱘的存活率和孵化率。
魚卵的黏性也有助于親魚護卵。一些魚類通過親魚的護卵行為以提高受精卵存活率,如大瀧六線魚、絨杜父魚等。大瀧六線魚由雄性選擇環境適宜受精卵孵化的水域筑巢,雌性將卵產在魚巢內,卵粒遇水硬化后,具有極強的黏性,牢固的黏附在基質上,這種繁殖策略有利于雄魚護卵,降低受精卵被捕食風險,提高其存活率[11]。
在無筑巢護卵習性的魚類中,卵的黏性有助于在整個胚胎發育過程中將魚卵固著在最佳孵化位置,對維持孵化率水平具有重要意義[12]。如歐洲白鮭Coregonuslavaretus既不護卵也不筑巢,卵粒和精子均撒播在選定的產卵場上,由于它們在水流湍急的水域產卵,所以幾乎不會存在卵沉積,受精卵孵化水環境中含氧量較高,有利于提高受精卵孵化率[13]。如果卵無黏性,受精卵可能會受到水流沖擊造成物理傷害,或被沖上岸邊因干燥而死亡,或被拖入含氧量較低、沉積物負荷較重的深水區,導致孵化率和存活率降低。有研究表明,歐洲白鮭受精卵死亡的主要原因是魚卵被水流從產卵場沖到不適合的發育區域[14]。
2 黏性卵的結構特征及其黏性機制
2.1 黏性卵的結構特征
大多數魚類的卵母細胞和卵被一層厚蛋白質膜所包覆[15],稱為卵膜。卵膜可以保護魚卵免受環境變化(如干燥、機械力)影響造成的器質性損傷[16-18],以及水流[19]、捕食者和病原體(如細菌和真菌)感染[20-21]的影響。卵膜可作為受精卵內部與外部環境污染物間隔離的屏障,在環境急劇變化時(如干燥)起到一定的保護作用,卵膜還有促進發育中胚胎氣體交換的功能[22]。
卵膜的結構包括一個較厚內層放射帶(zona radiata interna,ZRI)和一個較薄外層放射帶(zona radiata externa,ZRE)[23]。大多數魚類的卵均具有這種通用結構,然而,卵膜的形態可以根據物種和環境表現出修正[24]。非黏性魚卵通常具有較薄的卵泡上皮細胞層和放射帶,而黏性魚卵則被較厚的卵泡上皮細胞層和相對較復雜的卵膜包覆[25]。
覆蓋卵的放射帶是一個復雜的、充滿孔道的細胞外結構,孔道中填充著卵母細胞微絨毛和卵泡細胞突觸。放射帶通常分成兩層,每層均具有不同的形態特征[26]。內層主要由蛋白質組成,碳水化合物較少,與哺乳動物透明帶相似,其在受精和胚胎發育過程中的功能在進化中得到了保留[27];外層由特定的大分子組成,除糖蛋白外,還可能含有羧酸和硫酸多糖及少量唾液酸[28],主要負責卵與水環境的相互作用。魚類產卵后,放射帶經歷了廣泛的分子修飾,最終形成了卵膜。卵膜結構反映了對不同環境條件的適應性,無黏性卵通常具有較薄的卵膜,外層結構較簡單;黏性卵具有較厚和更復雜的卵膜,放射帶外層的附屬結構遇水會產生物理和化學變化,使其能夠黏附在不同的基質上[29]。
Rizzo等[1]對23個脂鯉目Cypriniformes和8個鲇形目Siluriformes魚卵進行了超微結構分析,并研究了卵的黏性及卵表面結構模式的相互關系,結果表明,脂鯉目魚類中,無黏性卵的放射帶表面呈孔道或纖絲網,弱黏性卵的放射帶表面僅為孔道,而高黏性卵的放射帶表面則為球狀、絲狀、絨毛狀或蜂窩狀的孔道;鲇形目魚類的卵表面結構均較相似,呈現常見的“膠狀層”結構,這一結構的魚卵是否具有黏性主要取決于膜卵表面的生化成分。
2.2 黏性卵的黏性機制
目前,對黏性魚卵表面的黏性物質成分及成因研究較少,據推測,卵表面黏性可能與生殖模式和系統發育有關[30],黏性魚卵的黏性機制研究目前也尚未完全清楚,現有的研究成果僅限具體試驗中魚類黏性機制的探討。黏性卵黏性機制的研究主要從形態學和生化兩方面入手,形態學方面運用電子顯微鏡對卵表面進行超微結構分析,研究卵表面特征性結構在遇水即呈現出黏性前后的變化;生化方面通常使用凝膠電泳法進行成分分析,探討卵膜表面大分子成分如糖蛋白等遇水化合后所產生的黏性物質成分及成因等。現有研究成果顯示,黏性卵的黏性機制根據魚的種類不同而有所差異。如香魚科和胡瓜魚科,包覆著魚卵的動物半球的黏性卵膜反轉,形成黏性膜[31-32];在雀鯛科Pomacentridae[33]、青鳉屬Oryziinae[34]、底鳉屬Fundulus[35]及蝦虎魚科Gobiidae[36],魚卵被卵膜上的長纖絲附著在基質上;另外,在很多種類的魚卵表面發現的黏性物質也使魚卵產生黏性,如鯡形目Clupeiformes、鯉形目Cypriniformes、鲇形目Siluriformes[37]、刺魚目Gasterosteiformes[38]、鱸亞目Percoidei[39]、鲉形目Scorpaeniformes[40]和鰈形目Pleuronectiformes[41]。
魚卵彼此黏附及魚卵黏附在基質上的方式隨環境條件[42]和魚卵表面結構的不同而變化。綜合分析,黏性的產生機制主要分為以下兩類。
2.2.1 形態學與生化雙重黏附機制 卵特殊的表面結構(如蜂巢結構、各種形態的突起、黏著絲等)與卵的黏度有重大關聯,這種特殊表面結構的魚卵通常具有形態學與生化雙重黏附機制。
研究表明,歐洲白鮭卵與水接觸30 s后開始產生黏性,且在與水接觸后幾分鐘內魚卵即完全黏附在一起[14]。對其卵膜超微結構觀察表明,與水接觸后,卵膜表面形態發生了顯著變化,絨毛狀突起附著在卵表面的結締絲上,通過這些突起的互相鎖定產生黏性,也可通過在卵表面與卵落入的基質間產生摩擦來促進黏附。這種絨毛狀突起與文鳊Vimbavimba卵的絨毛狀突起相類似,不過文鳊卵的黏附結構長度為4 μm,而歐洲白鮭卵上的突起更短且長度不一(1~3 μm)[43]。生化機制方面的分析顯示,與水接觸后的卵膜上存在一種水解蛋白酶(Chain A,RNase ZF-3e),而未與水接觸的卵則未發現該種水解蛋白酶的存在。歐洲白鮭卵的卵膜組織學檢測顯示,阿爾新藍和PAS染色反應均呈陽性,印證了糖蛋白及中性和酸性黏多糖的存在,此結果與在其他種類硬骨魚卵外部放射帶中發現的結果一致[19]。由此可見,歐洲白鮭卵的黏性源自卵膜,具有形態學和生物化學雙重黏附機制。
2.2.2 生化黏附機制 有些魚類具有相對平滑的卵表面結構(如放射帶孔道和膠狀外套層),此類魚卵是否具有黏性及黏性強弱則與具體魚的種類有關。此類卵黏性機制主要是通過生化機制實現黏附,即卵表面的化學物質,以及排卵后這些化學物質與水發生化合反應后產生的黏性物質,使魚卵黏附在基質上或相互黏附。
在有些魚類中,魚卵的黏性由外層放射帶ZRE的糖蛋白介導[43]。如鯉Cyprinuscarpio卵的黏性是蛋白質通過外層放射帶ZRE從卵黃周隙擴散而引起的[44]。在許多魚類中,黏性層來自卵泡上皮細胞,如歐鲇Silurusglanis,在卵黃生成作用中,卵泡上皮細胞轉化成結構不規則厚實的由黏性卵泡細胞組成的膠狀外套層。該膠狀外套層由黏多糖和黏蛋白組成,遇水膨脹成為飽滿富有彈性的黏性層,將魚卵附著在基質或魚巢中。這些黏性物質可以被蛋白酶分解,這表明這些黏性物質的基本成分是蛋白質[45]。
鱘魚卵的黏性成因仍然知之甚少。鱘魚卵受精后吸收水分形成卵黃周隙空間,從而引發卵膜膨脹并導致卵黏附[46-48]。卵泡上皮細胞的分泌物轉化形成了黏性層,這一層被認為是另一個卵泡上皮細胞層,但其更接近于放射帶。目前,這一黏性外層的存在已在多個鱘類中得到證實[49-53]。黏性層的主要成分是含有唾液酸的相對分子質量為110 000的糖蛋白 ,因此,使用蛋白水解酶可以有效地分解黏性層中的蛋白質。不同種的鱘類形成的黏性層厚度也有所不同[54],高首鱘Acipensertransmontanus的黏性層厚度為0.30~0.64 μm,西伯利亞鱘Acipenserbaerii為0.5~0.9 μm[55]。當卵子遇水后,外層在水中非生物因子的影響下產生黏性,如滲透壓、pH、溫度和離子含量(NaCl、KCl、CaCl2、Mg2SO4)等外部因素均會影響魚卵黏附的分子機制。
3 產黏性卵魚類人工授精及孵化技術
魚類人工繁育成功與否的主要因素是能否獲得高質量的受精卵。產黏性卵的魚類,由于卵的黏性特質導致在人工授精和孵化過程中存在較多困難,例如,卵膜表層結構遇水反應導致黏性卵在短時間內失去受精能力,受精卵黏連結塊致使內部氣體交換困難,進一步導致孵化率下降等。因此,產黏性卵的經濟魚類人工授精及孵化方法是生產過程中延續至今的研究課題,主要研究思路為兩個方向: ① 黏性魚卵的脫黏授精孵化法,使用簡便易操作的物理或化學方法使受精卵失去黏性,如利用黃泥漿水進行機械脫黏和利用水解蛋白酶進行化學脫黏,脫黏后再進行受精卵的孵化;② 人工魚巢接卵孵化法,此方法模擬自然環境中產黏性卵魚類的孵化條件,將受精卵接卵并附著在人工魚巢上進行孵化。常用的有棕櫚、水草為代表的傳統材料魚巢,聚乙烯網片為代表的新材料魚巢等。
3.1 脫黏孵化法
脫黏法是在人工催產的基礎上,先授精后脫黏,之后流水孵化的技術,此方法有效解決了黏性魚卵人工授精及孵化過程中的一些困難,具有受精率高、孵化管理方便、孵化效果好而穩定的優點,可以成批獲得規格整齊的魚苗。脫黏法大致可分為物理脫黏法和化學脫黏法。
3.1.1 物理脫黏法 物理脫黏主要利用脫黏劑對魚卵進行處理,例如黃泥漿、滑石粉、淀粉或富勒氏土等,使魚卵脫去黏性。亦可使用機械裝置對黏性卵進行沖洗分離魚卵,通過機械作用除去魚卵表面的黏液使魚卵脫去黏性。其中,最具代表性的方法是黃泥漿脫黏法,即采用黃泥漿與受精卵攪拌使之脫黏,然后洗去泥漿進行孵化。羅俊烈[56]采用鯽Carassiusauratus、鯪Cirrhinusmolitorella人工雜交后利用黃泥漿進行脫黏,然后流水孵化,此方法受精率可達92%,孵化率可達86%。 Monaco等[57]使用黃泥漿人工去除了高首鱘魚卵黏性,孵化率為54%~57%。Chebanov等[58]認為,富勒氏土是最有效的物質之一,但缺點是處理時間過長,使用黃泥漿和富勒氏土處理需要35~45 min。
陳力等[59]對白斑狗魚Esoxlucius的受精卵進行脫黏試驗,并使用6種材質的脫黏劑進行對比,分別為黃泥漿、滑石粉、生粉(淀粉)、牛奶、生豆漿和熟豆漿,對親魚人工催產授精后加入脫黏劑,結果表明,滑石粉脫黏效果最好,在顯微鏡下可觀察到卵的表面有大量分布均勻的小顆粒,適于規模化人工授精生產。郭長江等[60]對達氏鰉Husodauricus受精卵用質量濃度為200 g/L的滑石粉溶液進行脫黏,再用清水清洗后轉入尤先科孵化器,取得了良好的效果。
物理脫黏法的原理是利用機械震蕩和摩擦去除卵表面的黏性物質,操作不當會對受精卵產生損傷,病原體借此大量滋生而導致孵化率降低。Monaco等[57]指出,真菌感染是湖鱘Acipenserfulvescens卵孵化率下降的關鍵。Bouchard等[61]用質量濃度50 mg/L 的緩沖碘液處理魚卵30 min,然后再分別使用尿素-氯化鈉-單寧酸溶液及富勒氏土脫黏,這樣先消毒再脫黏的方法,雖然耗時增加,但孵化率高達99.6%和97.7%。
3.1.2 化學脫黏法 化學脫黏法是通過使用對魚卵卵膜產生化學作用的物質處理來去除魚卵的黏性,使用的材料包括蛋白水解酶、尿素-氯化鈉混合溶液和單寧酸等。
對于歐鲇和丁鱥Tincatinca,最有效地去除魚卵黏性的方法是使用蛋白水解酶。硬骨魚類的卵被一層厚的蛋白質層包覆,可通過蛋白水解酶分解蛋白質層去除黏性。傳統的物理脫黏劑(如黃泥漿、滑石粉和牛奶)在孵化期間通常會招致細菌和真菌的大量滋生。使用蛋白水解酶可以降低細菌和真菌感染的風險[62]。消除魚卵黏性的常用酶為堿性蛋白酶、麥芽糖酶、胰蛋白酶和α-胰凝乳蛋白酶。目前,堿性蛋白酶已成功用于去除鯉、丁鱥和歐鲇魚卵的黏性,該種酶在歐洲的許多孵化場中作為常規材料使用[63]。以鯉為試驗對象的研究中曾經使用過α-胰凝乳蛋白酶,但其效率(孵化率為50%~70%)比堿性蛋白酶(孵化率為80%~87%)低。麥芽糖酶曾被運用在尖齒胡鲇Clariasgariepinus卵脫黏處理中,僅需處理90 s,孵化率即可達71.6%[64]。此外,經6 IU/ mL的胰蛋白酶處理的里海鱘Acipenserpersicus卵,孵化率為86.9%[62]。研究表明,大多數酶處理通常需要1~2 min,李志等[65]對異育銀鯽Carassiusauratusgibelio的研究表明,在23 ℃時用質量分數為0.25%胰蛋白酶(pH為7.1~7.4)消化4 min可獲得脫黏受精卵。
尿素和NaCl的混合溶液是水產養殖中去除魚卵黏性最常用的方法[66]。Kowtal等[67]將高首鱘卵在質量分數為0.4%的尿素和質量分數為0.3%的NaCl溶液中處理35 min后,浸入質量分數為0.1%的單寧酸10 s,獲得的孵化率為57.7%,與黃泥漿處理組無明顯差異。比較尿素-氯化鈉-單寧酸溶液和富勒氏土處理湖鱘卵,結果孵化率分別為84.2%和85.6%[61]。
使用化學方法處理魚卵時,使用劑量和處理時間對其成功至關重要。不正確的劑量和過長的處理時間會增加死亡率[68]。浸入質量分數為0.15%的單寧酸溶液中60~90 s,丁鱥胚胎死亡率達65%~75%,而仔魚死亡率則高達85%~95%[69]。

3.2 人工魚巢接卵孵化法
采用傳統方法制作的黏性魚卵附著魚巢材料主要有棕櫚皮、水草(聚草、金魚藻)、楊柳根須等。蘇建國等[71]在人工繁育團頭魴的試驗中,利用楊樹根、棕櫚皮制作魚巢,孵化率為20%左右。楊雪軍等[72〗研究了黃顙魚對不同人工魚巢的產卵偏好,發現半球形棕櫚集卵效果優于半球形塑料和尼龍漁網制作的魚巢。張勝利[73]研究了用3種材料(芒萁片、棕櫚片、聚乙烯纖維)制作魚巢對錦鯉的集卵效果,發現芒萁制作的魚巢集卵效果最佳。
由于水草易腐爛,楊柳根須和棕櫚皮使用前需經水煮曬干,以除去單寧酸等有毒物質,其應用受到一定限制。刑克智等[74]采用帶框聚乙烯網片,網目為0.55~1.65 mm,網片先平鋪于布卵池,將受精卵布于網片上,待受精卵在網片上黏結牢固后,放入孵化池進行孵化,總孵化率可達80%。郭文等[75]采用相似的聚乙烯網片作為魚巢,將未受精的魚卵整形成單層且緊密排列的卵片后,再進行授精,避免了魚卵變成團塊狀,使魚卵受精時能與精子充分接觸,提高了魚卵受精率。
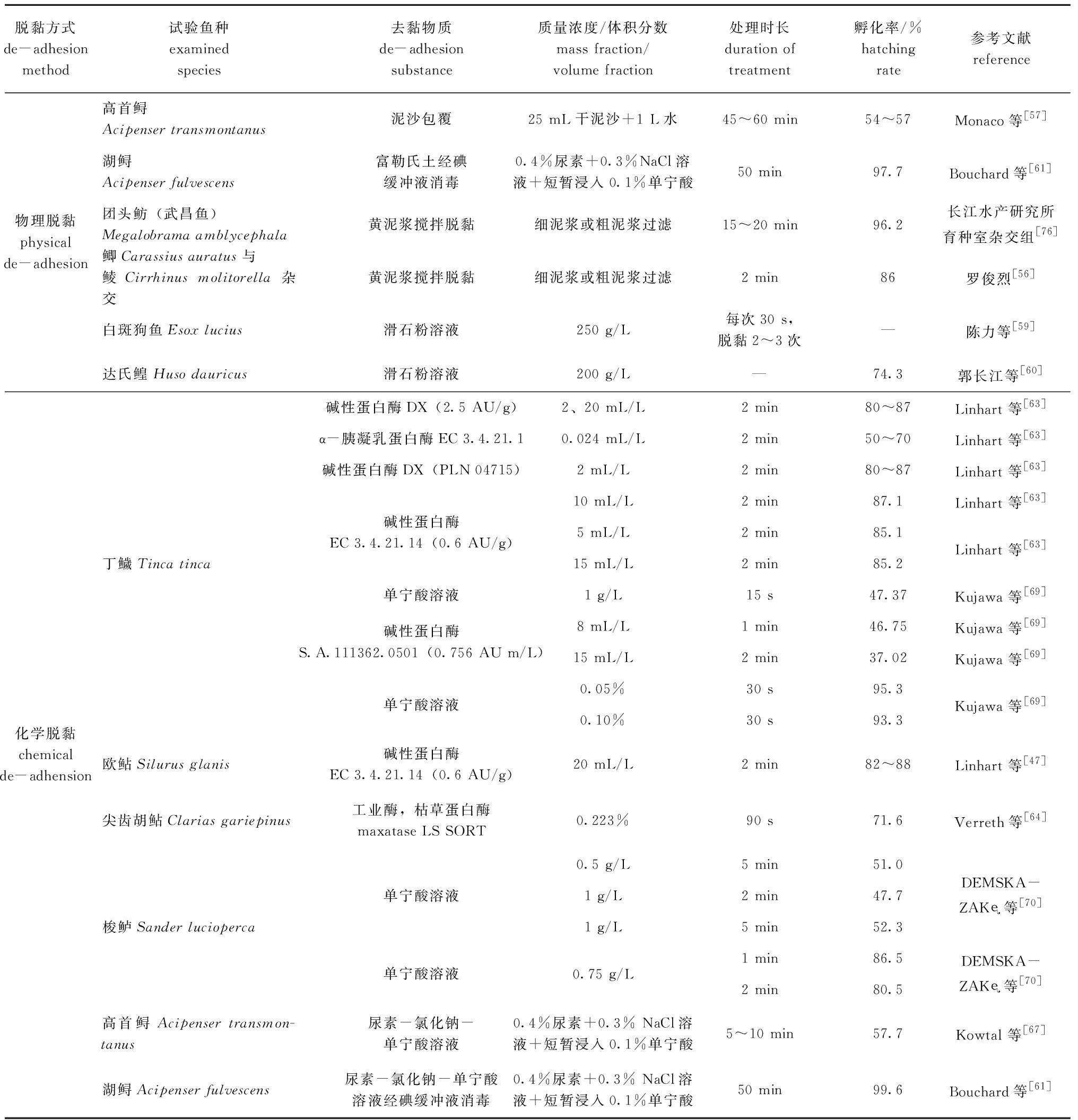
表1 常見養殖魚類黏性卵脫黏方法及孵化率Tab.1 De-adhesion methods of adhesive eggs and corresponding hatching rates in common fishes
4 存在問題及展望
抑制魚卵黏性的活化,或使已經與水化合發生黏附的魚卵脫黏,是黏性卵人工繁育技術革新最重要的研究方向之一。近年來,在國內外科研人員的不斷努力下,被作為研究對象的魚類愈加廣泛,對黏性卵的研究也日漸深入,但仍存在一定局限和不足。本研究中綜合國內外學者的最新研究進展,在黏性卵研究內容較為薄弱方面提出了幾點尚需加強和拓展的研究內容,以期為今后的研究提供參考。
1) 建立黏性卵黏附程度的統一量化評估標準。目前只能憑借肉眼簡單鑒別非黏性卵、弱黏性卵和黏性卵,實際上,不同魚類黏性卵的黏性程度存在較大差異。卵黏性程度不同,意味著黏性卵的表面超微結構、表面黏性層成分、黏性機制有差異,生產實踐中對卵黏性的抑制及針對性脫黏處理方法也不同。因此,建立黏性卵黏附程度的統一量化評估標準是黏性卵研究的基礎之一,可以作為今后的一個基礎性研究方向。
2) 加強對黏性卵表面結構特別是超微結構的研究。目前,黏性卵表面結構的分類系統尚無學術界較為公認的體系。使用先進的電子顯微鏡技術對黏性卵做深入的超微結構研究,建立各魚類魚卵形態的“數據庫”,完善黏性卵表面結構分類體系,這對魚類的辨識和分類,以及了解魚類生活的環境因素十分重要。這些工作不僅僅是基礎性的學科研究,對提高人工繁育效率和改進工廠化養殖技術也具有重大指導意義。
3) 加強黏性卵表面黏性成分的研究。黏性卵通常具有形態與生化雙重黏附機制,目前僅對少數魚類黏性卵表面黏性成分進行了詳盡的描述,且這些黏性蛋白成分的成因,遇水化合反應后對卵表面結構變化的影響等,現有研究成果還有較大的延展空間,可以作為重要的研究方向。特別是對黏性卵表面蛋白質和黏多糖成分的研究,可以指導生產實踐,有針對性地使用蛋白酶分解的方法,對魚卵進行高效且低成本的脫黏處理。
4) 脫黏技術的改進。現有關于黏性卵魚類的人工繁育孵化技術中,最關鍵的環節之一是對黏性卵的脫黏。實際生產中還可以考慮使用配方溶液,在黏性卵發生黏附之前抑制黏附,這可能是一種更有效的方法。
