兒童下肢長骨干骨折后過度生長的影響因素及機制研究進展
2021-03-04 05:20:12岳宇航綜述李連永審校
臨床小兒外科雜志 2021年2期
岳宇航 綜述 李連永 審校
下肢長骨干骨折是兒童常見創傷,主要包括股骨干骨折和脛骨干骨折,治療方式包括保守治療和手術治療。 但無論采用何種治療方式,雙下肢不等長是兒童下肢長骨干骨折后最常見的并發癥之一,通常為患肢過度生長所致[1]。 雙下肢不等長除了引起跛行,還可引起背痛、脊柱側彎以及坐骨神經痛等癥狀。 輕微下肢不等長的患者可以通過墊高鞋跟的方法矯正,較為嚴重的患者需行臨時骨骺阻滯術或肢體延長術[2]。 下肢長骨骨折后肢體過長的發生機制尚不完全明確,本文對其研究進展進行綜述。
一、骨折后過度生長現象
1849 年,有學者發現兒童長骨干骨折后會出現過度生長的現象,隨后Paget(1863 年)、Ollier(1867年)等也相繼報道了類似現象。 1921 年,Truesdell詳細記錄了5 例股骨干骨折兒童的受傷部位、治療方式以及隨訪過程,發現患側下肢存在過度生長的情況,并認為創傷本身是引起股骨加速生長的主要原因[3]。
隨著現代影像學技術的發展,對這一現象的認識也越來越明確。 相關臨床研究表明,過度生長主要發生于骨折后的18 ~24 個月,多見于12 歲以內的患者,7 歲以內患者更為明顯,雙下肢長度差通常在10 mm 左右[1,4,5]。 股骨干骨折的患者,不論是采用閉合復位石膏固定或Pavlik 吊帶治療,還是切開復位內固定,均有發生過度生長的現象[4]。 Flynn等[6]對過度生長現象提出了評價指標,認為過度生長造成雙下肢長度差在1 cm 以內的患者預后優秀,1 ~2 cm 者預后滿意,2 cm 以上者預后較差。
對于引起過度生長的危險因素,既往研究普遍集中在性別、年齡、骨折原因、骨折類型、骨折線位置等方面。 Clement 和Colton 通過分析比較44 例年齡在7 個月至12 歲經保守治療的股骨干骨折患者,發現男孩平均過度生長9.9 mm,女孩平均過度生長5.5 mm(P<0.025)[7]。 但他們并沒有分析不同性別引起過度生長差異的具體原因,而有學者認為過度生長與性別無關[8,9]。 同時,該研究也注意到年齡越小過度生長越明顯,與其他學者研究結果一致[10-12]。
關于骨折部位,Beaty 等[13]認為骨折線位于股骨近端1/3 處對過度生長的影響更為明顯。 而Edvardson、Syversen 以及Shapiro[11,14]并未發現上述現象。 骨折類型也被認為是影響過度生長的因素。國內有研究認為不穩定型,即長斜形、旋轉和粉碎性股骨干骨折較穩定型(短斜形和橫形)股骨干骨折更易出現過度生長超過1 cm 的現象[15]。 Fakoor等[8]也發現在同樣采用切開復位鋼板螺釘固定的情況下,股骨干螺旋形骨折較橫形骨折患者過度生長更明顯。 該現象可能與不穩定骨折的穩定性較差及出血較多有關。 由此可見,過度生長現象發生的危險因素仍存在廣泛的爭議,需要更為全面的臨床研究證實。
內固定物的選擇也可能與骨折后過度生長有關。 近年來,彈性髓內釘廣泛應用于兒童長骨骨折,其與肢體過長的相關性也備受關注。 有研究發現兒童脛骨干骨折彈性髓內釘固定2 年后,隨訪時患側可平均過度生長5 mm,最長可達15 mm[16]。Park 等[9]通過分析股骨干骨折患者彈性髓內釘與髓腔直徑的比例,發現比例≥0.8,股骨平均過度生長0.36 cm,而<0.8 時,過度生長可達1.0 cm,兩者有統計學差異,國內相關研究也支持這一觀點[15]。這一差異可能與髓內釘占髓腔比例越高,骨折固定越穩定有關[17]。 Nascimento 等[18]通過比較彈性髓內釘與手法復位石膏固定治療股骨干骨折的結果,發現彈性髓內釘治療更易導致股骨過長,該結果可能與手法復位石膏固定更易出現成角畸形有關。但這些研究并未分析過度生長與是否解剖復位有關系。 尚無研究分析彈性髓內釘固定方式(雙側鉆孔CC 型和單側鉆孔CS 型)在過度生長方面有無差別。 另外,彈性髓內釘導致骨折后過度生長的機制尚不明確,仍缺乏不同內固定方式對過度生長影響的比較研究。 對于不伴有其他損傷的脛骨干骨折,尚無研究表明其脛骨過度生長是否伴隨腓骨過度生長,以及是否產生踝外翻畸形。 值得注意的是,一些學者在上肢長骨骨折及下頜骨折患者中亦觀察到過度生長的現象,其發生機制及危險因素是否與下肢長骨骨折相似有待進一步研究[19,20]。
二、骨膜斷裂與骨折后過度生長
既往研究普遍認為骨折后過度生長與骨折后干骺端血供增加以及骨膜完整性被破壞有關。 Yabsley 和Harris[21]發現長骨骨干骨折以及骨膜剝離后生長板的血運有所增加。 而更多研究集中在骨膜受損與過度生長的關系。
早在1873 年,Ollier[22]發現骨折后過度生長與骨膜的完整性受損有關,并制作了相關動物模型。該研究將尺橈骨末端的骨膜剝離,發現骨骼生長速度增快。 隨后關于骨膜與過度生長的研究逐漸展開。 Halanski 等[23]通過橫形切開、剝離兔一側脛骨近端骨膜,發現2 ~8 周后脛骨遠端生長板細胞增殖分化較對側活躍。 與此同時,該研究測量了2 周和8 周時脛骨近端生長速率,發現2 周時術側較對側快(64.1 ±29)μm/d(P<0.001);8 周時術側生長較快但無統計學意義(P=0.75)。 Warrell 等[24]通過比較不同方向切開骨膜致過度生長情況,發現環形切開骨膜較縱向切開骨膜更易導致過度生長,因此推測沿長骨干縱軸方向的牽引力降低是促進骨骼過度生長的原因。 為證實上述觀點,Morscher等[25]利用外固定架,對脛骨骨膜橫形切開后的兔子在脛骨近端生長板兩側施加或降低1 kg 的壓力,研究骨膜斷裂后生長板壓力之間的關系。 實驗證明,骨膜斷裂后的確會降低生長板兩端的壓力,而導致骨骼過度生長。
對于生長板兩端壓力與骨生長的關系,Hueter-Volkmann 定律指出,生長板所受壓力增加,骨的生長就會受到抑制;相反,骨的生長就會加速[26]。Strokes 等[26]在動物研究中發現,不論是椎骨還是脛骨,在一定范圍內,生長板兩端壓力降低與骨骼長度增加呈線性關系。 相關動物實驗結果對將來臨床治療雙下肢不等長可能有一定的指導意義。 Limpaphayom 和Prasongchin[27]曾嘗試通過骨膜剝離治療雙下肢不等長患者,并取得較好效果。
骨膜完整性受損與生長板過度生長的關系可以從生物力學角度加以解釋。 骨膜是一種纖維結締組織,在骨表面形成一個連續的鞘。 纖維骨膜主要附著于骨兩端的干骺端和生長板,其在生長板兩端產生的壓力可能會對骨骼發育造成影響。 Bertram 等[28]通過測量不同位置骨膜張力,繪制了張力應變曲線,并發現骨膜張力從生長板近端到生長板遠端每毫米增加1 牛頓。 根據張力應變曲線,張力變化越快,即骨膜粘附越緊的地方,兩端產生的壓力就越大,因而生長板和干骺端是壓力最大的地方。 骨膜橫形切斷之后,骨膜的張力減小或消失,從而導致生長板兩端的壓力減小,骨生長加速。 這可以推測,粉碎性骨折或螺旋形骨折后的股骨遠端生長板可能因骨膜破壞更嚴重,兩端壓力減低更多,故此類型的骨折較其他穩定性骨折有更加明顯的過度生長現象。
上述實驗均從一個較為宏觀的角度對骨折后過度生長現象進行分析和解釋,沒有探究相關的分子生物學機制,未能揭示過度生長的本質過程。
三、長骨骨折后過度生長相關分子機制
兒童長骨的生長主要由兩端的生長板完成,但不同骨骼遠近端骺板對生長的貢獻不同。 股骨近端生長板對股骨生長貢獻較小,約占股骨總長度的30%;股骨遠端生長板呈盤狀,承擔了約70%的生長任務;脛骨近端和遠端生長板對脛骨的貢獻分別為55%和45%[2]。 Bisgard[29]于1936 年在動物實驗中發現骨折后過度生長現象集中在生長板,而不是骨折部位。 作者在臨床中也發現,骨折后患側Harris 線與骺板的距離較對側更寬(圖1),說明骨折后過度生長現象來自于骺板。
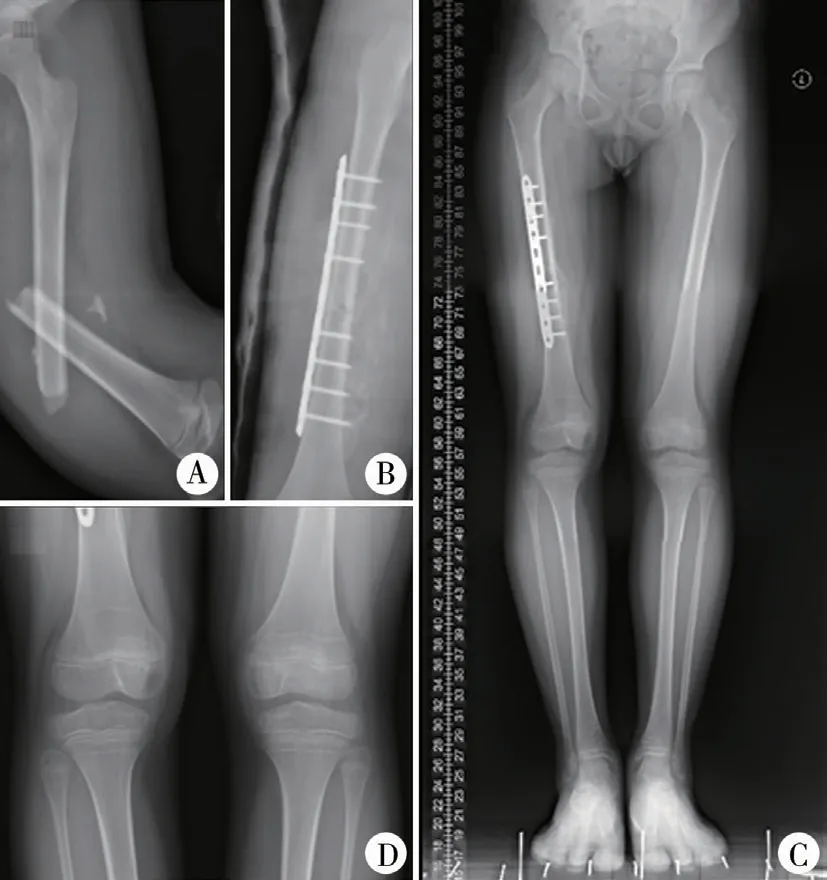
圖1 股骨干骨折后過度生長 注 患者,男,8 歲,車禍傷致右股骨中段骨折,行切開復位鋼板內固定治療,術后1 年復查 A:右股骨粉碎性骨折術前; B:右側股骨鋼板螺釘內固定術后; C:右側全長728.4 mm,左側全長709.0 mm; D:右側Harris 線距離生長板上緣寬度大于左側Fig.1 Overgrowth after femoral shaft fracture
生長板是一種特殊的軟骨結構,依據組織學和功能特點可以分為靜止層、增殖層、肥大層和鈣化層。 軟骨細胞沿軸線方向增殖,按柱狀排列,并在肥大層中體積增大。 靜止細胞和肥大細胞附近膠原也逐漸縱向排列,與柱狀排列的細胞走行一致。生長板發育與一些全身性激素以及旁分泌因子有關,主要包括性別決定區Y 框蛋白9(SRY-box transcription factor 9,Sox9)、Runt 相關轉錄因子2(Runtrelated transcription factor 2,Runx2)、Wnt 家族、印猬蛋白(Indian Hedgehog,IHH)、甲狀旁腺激素相關蛋白(parathyroid hormone related protein,PTHrP)等特異性轉錄因子;成纖維細胞生長因子(fibroblast growth factor,FGF)家族,以骨形態發生蛋白(bone morphogenetic protein,BMP)為代表的轉化生長因子-β(transforming growth factor-β,TGF-β)超家族,血管內皮生長因子(vascular endothelial growth factor,VEGF)等以及甲狀腺激素、性激素等[30]。 Smad1 可以促進BMP2 誘導骨髓間充質干細胞向軟骨細胞轉化,同時抑制成骨細胞分化[31]。 Sox9 在軟骨形成中起到關鍵作用,在胎兒時期即有表達,主要促進骨祖細胞向軟骨細胞轉化[32]。 同時,Sox9 也與性別決定因子SRY 有密切關系[33],或可以解釋Clement等[7]觀察到男孩過度生長較女孩明顯的現象。Runx2 在軟骨成熟過程中發揮重要作用,通過Runx2-Ihh-PTHrP 通路負反饋調節生長板軟骨細胞肥大,此外IHH 也可以在軟骨膜中刺激Runx2 表達,在成骨細胞分化中發揮作用[34]。 FGF 可以與跨膜蛋白成纖維細胞生長因子受體(fibroblast growth factor receptor,FGFR)胞外區域結合,誘導其胞內區域酪氨酸殘基磷酸化。 活化的FGFR 可以在胞內募集相關靶蛋白并將它們磷酸化修飾[35]。 FGF 下游通路包括JAK/STAT、MAPK、PI3K/AKT、PLCγ/PKC等,主要負責調節生長板軟骨細胞增殖、分化、存活以及產生胞外基質[36]。 FGF 家族和BMP 家族可以相互調節各自下游通路的表達以及它們的穩定性[37]。 上述因子共同作用,促進生長板軟骨向骨轉化并使長骨延長,完成軟骨內化骨。
骨折后過度生長的分子機制可能與上述生物學通路有關。 Ashraf 等[38]使用基因芯片測定大鼠在股骨干骨折后股骨近端生長板中相關基因的mRNA 表達,發現血管再生相關基因表達下調,這與Warrell 在動物模型中發現干骺端血供增加相沖突[24]。 與神經細胞相關的轉錄因子Marcks 樣蛋白表達增多,促進4,5-二磷酸磷脂酰肌醇(phosphatidylinositol 4,5-bisphosphate,PIP2)轉化為三磷酸肌醇(inositol 1,4,5-trisphosphate,IP3)、二酰甘油(diacylglycerol,DAG)、3,4,5-三磷酸磷脂酰肌醇(phosphatidylinositol 3,4,5-trisphosphate,PIP3)等信號分子,激活多條信號通路,刺激長骨軸向生長有關[39]。與巨噬細胞、破骨細胞相關的基因表達增加,說明骨轉換的速度增快。 與此同時,成骨細胞和骨細胞相關基因表達也出現上調,骨形成過程同樣加快。與預想的一樣,細胞周期相關蛋白的表達均有所增加。 細胞周期特異性蛋白1 和tramdorin 1 蛋白在骨折后的一段時間內表達均上調,表明細胞增殖活躍。 除此之外,Fischerauer 等[40]研究發現BMP-6 和BMPR-1a 在脛骨骨折的動物模型中表達也有所增加,說明骨髓間充質干細胞向軟骨細胞分化增加。另外,相關基礎研究發現,FGFR3 可以通過一種蛋白酶依賴的方式調節BMPR-1a 表達[41],故考慮FGF 家族可能也參與了骨折后過度生長。
這些實驗研究了骨折后過度生長的分子機制,但仍有不足。 首先,股骨干骨折后股骨遠端生長板生長的長度明顯多于股骨近端。 而Ashraf 等[38]采用生長較少的近端,其代表性可能存在欠缺。 其次,缺少原位雜交和免疫組化的研究,同時未能揭示轉錄后蛋白質水平的調控。 既往實驗也未能從分子生物學角度解釋骨膜破壞、生長板兩端壓力降低對生長板過度生長的影響,故仍需進一步研究。
四、問題與展望
兒童下肢長骨干骨折后過度生長一直是困擾小兒骨科醫生的一個難題。 過度生長可能給患者家庭帶來更大的經濟負擔。 臨床上尚無法確定過度生長現象與哪些因素有關,也無明確方案減少或避免該并發癥的發生。 目前只能通過定期復查,盡早發現并及時對癥治療。 研究兒童下肢長骨干骨折后過度生長的危險因素有利于提高患者預后及生活質量,符合小兒骨科醫生和患者家屬的期盼,也有助于減少社會醫療負擔。
隨著科技的進步,對于過度生長的研究進入分子生物學領域,但其分子機制的研究仍然較少。 過度生長現象可能涉及到復雜的分子生物學機制,多條通路共同調控,但骨折是其始動因素。 相關基因的過表達,細胞因子的刺激等方面還有待進一步研究。 另外尚不清楚生長板壓力降低后生長加速的原因,該現象可能與某種壓力感受器傳遞相關信息、激活分子通路、刺激細胞增殖分化有關。 后續實驗可以利用基因組學、轉錄組學、蛋白質組學等方法探尋相關蛋白表達的差異,研究相應蛋白通路在過度生長現象中的作用。
認識兒童下肢長骨干骨折后過度生長這一現象的分子生物學機制,有利于臨床醫生對下肢長骨干骨折后過度生長的早期認識及預防,并可利用這一機制治療先天性或后天性雙下肢不等長。 如何實現對過度生長這一現象的控制并將其運用到臨床,是需要攻克的一個難題。 綜上所述,兒童下肢長骨干骨折后過度生長現象在病因上仍無明確結論,其相關分子機制也較為復雜且研究較少,仍需臨床醫生及基礎研究學者不斷攻堅克難,進一步深入探索。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
