斑節(jié)對(duì)蝦3個(gè)種質(zhì)群體體質(zhì)量性狀配合力及雜種優(yōu)勢(shì)分析
2021-02-28 03:15:58周發(fā)林楊其彬楊麗詩李運(yùn)東黃建華江世貴
南方水產(chǎn)科學(xué) 2021年1期
關(guān)鍵詞:質(zhì)量
周發(fā)林 ,楊其彬 ,姜 松 ,楊麗詩,李運(yùn)東,黃建華,江世貴
(1.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所熱帶水產(chǎn)研究開發(fā)中心,海南 三亞 572018;2.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所/農(nóng)業(yè)農(nóng)村部南海漁業(yè)資源開發(fā)利用重點(diǎn)實(shí)驗(yàn)室,廣東 廣州 510300;3.中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所深圳試驗(yàn)基地,廣東 深圳 518121)
斑節(jié)對(duì)蝦 (Penaeus monodon) 肉質(zhì)鮮美、營(yíng)養(yǎng)豐富、經(jīng)濟(jì)價(jià)值高,是體形最大的對(duì)蝦,也是我國(guó)非常重要的對(duì)蝦養(yǎng)殖品種之一,非常適合大規(guī)格養(yǎng)殖[1]。大規(guī)格蝦價(jià)格高、效益好,國(guó)內(nèi)外市場(chǎng)需求量大,但是最近十多年,由于斑節(jié)對(duì)蝦親蝦種質(zhì)質(zhì)量良莠不齊,繁育的苗種質(zhì)量無法得到保障,限制了我國(guó)斑節(jié)對(duì)蝦養(yǎng)殖產(chǎn)業(yè)的發(fā)展,其養(yǎng)殖產(chǎn)量一直在7 萬~10 萬噸徘徊,因此,開展斑節(jié)對(duì)蝦遺傳育種工作,培育出具有生長(zhǎng)快速等優(yōu)良經(jīng)濟(jì)性狀的新品種,是斑節(jié)對(duì)蝦養(yǎng)殖產(chǎn)業(yè)亟需解決的問題。2005 年,中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所在攻克了斑節(jié)對(duì)蝦親蝦規(guī)模化全人工培育技術(shù)難題的基礎(chǔ)上,啟動(dòng)了斑節(jié)對(duì)蝦育種研究,經(jīng)過多年努力,建立了斑節(jié)對(duì)蝦育種技術(shù)體系,并培育獲得斑節(jié)對(duì)蝦“南海1 號(hào)”新品種[2]和多個(gè)抗逆新品系。不過由于缺乏良種,制約其產(chǎn)業(yè)發(fā)展的基本格局仍然沒有得到根本性改變,其主要原因是我國(guó)養(yǎng)殖區(qū)域環(huán)境差異大,對(duì)斑節(jié)對(duì)蝦品種多樣化要求高,已有斑節(jié)對(duì)蝦良種不能完全滿足其養(yǎng)殖業(yè)的需求,因此,斑節(jié)對(duì)蝦育種是一項(xiàng)長(zhǎng)期且不能間斷的工作。目前,斑節(jié)對(duì)蝦良種培育仍然以選擇育種為主。為了獲得更多具有不同優(yōu)良經(jīng)濟(jì)性狀的良種,開展斑節(jié)對(duì)蝦選育技術(shù)的創(chuàng)新十分必要。
雜交育種是改良水產(chǎn)動(dòng)物重要經(jīng)濟(jì)性狀、培育優(yōu)良品種的有效手段之一[3-4]。在水產(chǎn)動(dòng)物種內(nèi)雜交育種研究中,配合力是非常重要的遺傳參數(shù),也是雜種優(yōu)勢(shì)利用的必要前提[5-6]。配合力分為一般配合力 (General combining ability,GCA) 和特殊配合力 (Specific combining ability,SCA),在玉米雜交育種中最早提出[7],反映不同品系或群體間的性狀配套效果,在農(nóng)作物中廣泛應(yīng)用[8-11]。在水產(chǎn)動(dòng)物中,配合力和雜交優(yōu)勢(shì)研究主要集中在魚類[12-15]和貝類[5,16-17],其他水產(chǎn)動(dòng)物也有報(bào)道,如刺參(Apostichopus japonicus) 等[18]。在對(duì)蝦中,關(guān)于凡納濱對(duì)蝦 (Litopenaeus vannamei) 不同群體配合力和雜種優(yōu)勢(shì)分析研究較多。王浩等[19]分析了凡納濱對(duì)蝦7 個(gè)引進(jìn)群體體質(zhì)量和存活性狀的配合力;胡志國(guó)等[20-21]分析了3 個(gè)凡納濱對(duì)蝦引進(jìn)群體對(duì)溫度、鹽度的耐受力以及耐氨氮和低溶氧的配合力。
不過到目前為止,還未見到關(guān)于斑節(jié)對(duì)蝦群體間的生長(zhǎng)等重要經(jīng)濟(jì)性狀的配合力分析。因此,本研究分析了斑節(jié)對(duì)蝦“南海1 號(hào)”、非洲選育品系和泰國(guó)群體之間的體質(zhì)量配合力和雜種優(yōu)勢(shì),以期為斑節(jié)對(duì)蝦雜交育種研究提供重要理論依據(jù)和技術(shù)指導(dǎo)。
1 材料與方法
1.1 親本來源
1) 斑節(jié)對(duì)蝦“南海1 號(hào)”(S):以我國(guó)海南三亞、臨高、文昌和泰國(guó)南部 (普吉島) 附近海域野生親蝦為初始親本,以體質(zhì)量性狀為選育目標(biāo),從2005 年開始,經(jīng)連續(xù)多代群體選育獲得,并在2010 年通過了全國(guó)水產(chǎn)原種和良種審定委員會(huì)審定。該品種的主要優(yōu)點(diǎn)在于同等條件下其生長(zhǎng)速度比未選育蝦提高20%以上。
2) 非洲品系 (A):從2009 年開始,以非洲南部附近海域斑節(jié)對(duì)蝦野生種質(zhì)資源群體為基礎(chǔ)群體,以體質(zhì)量生長(zhǎng)速度和耐高氨氮為選育指標(biāo),經(jīng)過多代的家系選育和BLUP 育種獲得的新品系。
3) 泰國(guó)群體 (T):2010 年,從泰國(guó)引進(jìn)的泰國(guó)野生資源種質(zhì)群體700 多尾,通過活體傳代保存的種質(zhì)資源群體。
以上3 個(gè)種質(zhì)群體保種于中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所深圳試驗(yàn)基地 (斑節(jié)對(duì)蝦遺傳育種中心)。
1.2 雙列雜交群體的構(gòu)建
2012 年3 月,在中國(guó)水產(chǎn)科學(xué)研究院南海水產(chǎn)研究所深圳試驗(yàn)基地 (斑節(jié)對(duì)蝦遺傳育種中心),利用非洲品系群體、“南海1 號(hào)”群體和泰國(guó)群體作為親本個(gè)體,開展了3 個(gè)斑節(jié)對(duì)蝦種質(zhì)群體內(nèi)自繁和群體間雜交試驗(yàn)。采用人工授精技術(shù),3 個(gè)群體親本依照完全雙列雜交的方式交配,構(gòu)建3 個(gè)自交群體和6 個(gè)雜交群體 (表1)。

表1 3個(gè)群體雙列雜交交配組合Table 1 Complete diallet cross of three populations
1.3 苗種培育與養(yǎng)殖
每個(gè)交配組合選取10~15 尾成熟時(shí)間相差不到3 d 的雌蝦。選取每尾雌蝦孵化出的無節(jié)幼體約6 000 尾,放入500 L 的培育桶單獨(dú)培育至仔蝦后,運(yùn)往中國(guó)水產(chǎn)科學(xué)研究院熱帶水產(chǎn)研究中心安游基地進(jìn)行標(biāo)粗培育。培養(yǎng)至約3 cm 時(shí),每個(gè)交配組合隨機(jī)選取800 尾個(gè)體進(jìn)行熒光標(biāo)記,以區(qū)分不同交配組合,然后把標(biāo)記好的所有交配組合個(gè)體混養(yǎng)于130 m2水泥池中。
1.4 數(shù)據(jù)測(cè)定
個(gè)體養(yǎng)殖至4 月齡時(shí),從每個(gè)群體中隨機(jī)抽出150 個(gè)個(gè)體用于體質(zhì)量測(cè)量。體質(zhì)量用電子天平稱量,精確到0.01 g。
1.5 數(shù)據(jù)處理
利用Excel 2013 進(jìn)行體質(zhì)量數(shù)據(jù)整理,使用NFN 9.3 進(jìn)行不同交配組合體質(zhì)量的方差分析和多重比較。配合力和雜種優(yōu)勢(shì)的計(jì)算按文獻(xiàn)方法[14,21-23]。
體質(zhì)量性狀的配合力分析模型為:
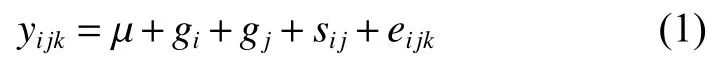
式中yijk為第i個(gè)父本群體與第j個(gè)母本群體雜交的第k個(gè)個(gè)體的體質(zhì)量數(shù)據(jù),μ為總體均值,gi(gj) 為第i(j) 個(gè)群體的父本 (母本) 一般配合力 (隨機(jī)效應(yīng)),sij為第i個(gè)父本群體與第j個(gè)母本群體雜交的特殊配合力 (隨機(jī)效應(yīng)),eijk為隨機(jī)誤差效應(yīng)。
按照下列公式計(jì)算雜種優(yōu)勢(shì):

式中 A 為所有雜交群體的體質(zhì)量均值,B 為所有自交群體的體質(zhì)量均值,F(xiàn)1為某一雜交組合的體質(zhì)量平均值,PM為雙親的體質(zhì)量平均值,PB為優(yōu)親的體質(zhì)量平均值。
2 結(jié)果與分析
2.1 3 個(gè)群體體質(zhì)量性狀的描述性統(tǒng)計(jì)、方差分析和多重比較
從每個(gè)交配組合中隨機(jī)選取150 個(gè)個(gè)體進(jìn)行體質(zhì)量收集工作,共獲得1 350 個(gè)數(shù)據(jù),各交配組合體質(zhì)量介于5.50~17.90 g,變異系數(shù)介于0.17~0.36。斑節(jié)對(duì)蝦各交配組合間體質(zhì)量存在一定的差異,其中AS 和SA 組合體質(zhì)量顯著高于3 個(gè)自交群體 (AA、SS 和TT) (表2 和圖1),研究結(jié)果表明6 個(gè)雜交組合的體質(zhì)量性狀遺傳改良潛力較大。
2.2 3 個(gè)群體之間的配合力分析
在水產(chǎn)動(dòng)物重要經(jīng)濟(jì)性狀的配合力分析中,廣泛采用了混合線性模型[12,19-21,24]。本研究也采用了混合線性模型,對(duì)斑節(jié)對(duì)蝦3 個(gè)不同來源種質(zhì)群體的體質(zhì)量性狀的配合力進(jìn)行了分析,為斑節(jié)對(duì)蝦體質(zhì)量性狀雜交優(yōu)勢(shì)利用和良種選育提供重要參考數(shù)據(jù)。
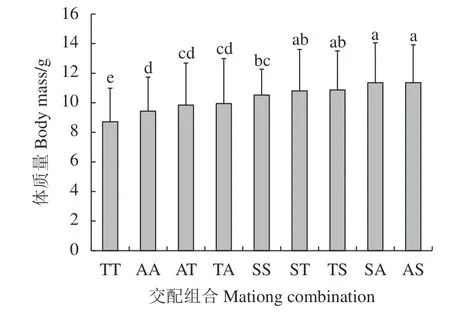
圖1 9個(gè)交配組合收獲體質(zhì)量的多重比較圖中不同小寫字母表示差異顯著 (P<0.05)Figure 1 Multiple comparison of harvest body mass of nine mating combinationsDifferent lowercases indicate significant difference (P<0.05).
2.2.1 3 個(gè)群體一般配合力 (GCA) 分析 斑節(jié)對(duì)蝦3 個(gè)群體體質(zhì)量性狀的一般配合力見表3。不同群體體質(zhì)量性狀一般配合力數(shù)值從大到小依次為:“南海1 號(hào)”群體>非洲品系群體>泰國(guó)群體,其數(shù)值分別為0.437 3、?0.062 4 和?0.374 9。“南海1號(hào)”群體的體質(zhì)量性狀一般配合力最高。

表3 3個(gè)群體體質(zhì)量性狀的一般配合力效應(yīng)值Table 3 General combining ability in harvest body mass of three populations
一般配合力是由基因的加性效應(yīng)所致[25-26],而遺傳力也是由加性效應(yīng)決定。因此,一個(gè)種群一般配合力高,初步預(yù)測(cè)其具有較好的選育潛力,作為親本能生產(chǎn)性狀優(yōu)良的子代。根據(jù)本研究數(shù)據(jù)分析,“南海1 號(hào)”群體的一般配合力最高且為正值,其次是非洲品系選育群體,最低為泰國(guó)群體,且2 個(gè)群體的一般配合力均為負(fù)值。研究結(jié)果表明,較非洲品系選育群體和泰國(guó)群體,“南海1 號(hào)”群體在質(zhì)量性狀方面存在較為豐富的加性遺傳變異。在斑節(jié)對(duì)蝦體質(zhì)量性狀方面良種選育時(shí),可以考慮該品種作為主要親本來源之一。
2.2.2 3 個(gè)群體特殊配合力分析 斑節(jié)對(duì)蝦9 個(gè)交配組合的體質(zhì)量性狀的特殊配合力 (SCA) 見表4。3 個(gè)自交群體 (AA、SS 和TT) 的體質(zhì)量特殊配合力均為負(fù)值,其數(shù)值分別為?0.676 6、?0.608 3 和?0.779 8。6 個(gè)雜交組合體質(zhì)量特殊配合力數(shù)值從大到小排序?yàn)锳S>SA>TS>ST>TA>AT,其中非洲品系選育群體ד南海1 號(hào)”群體 (♀)(AS) 體質(zhì)量的特殊配合力最高,其效應(yīng)值為0.613。
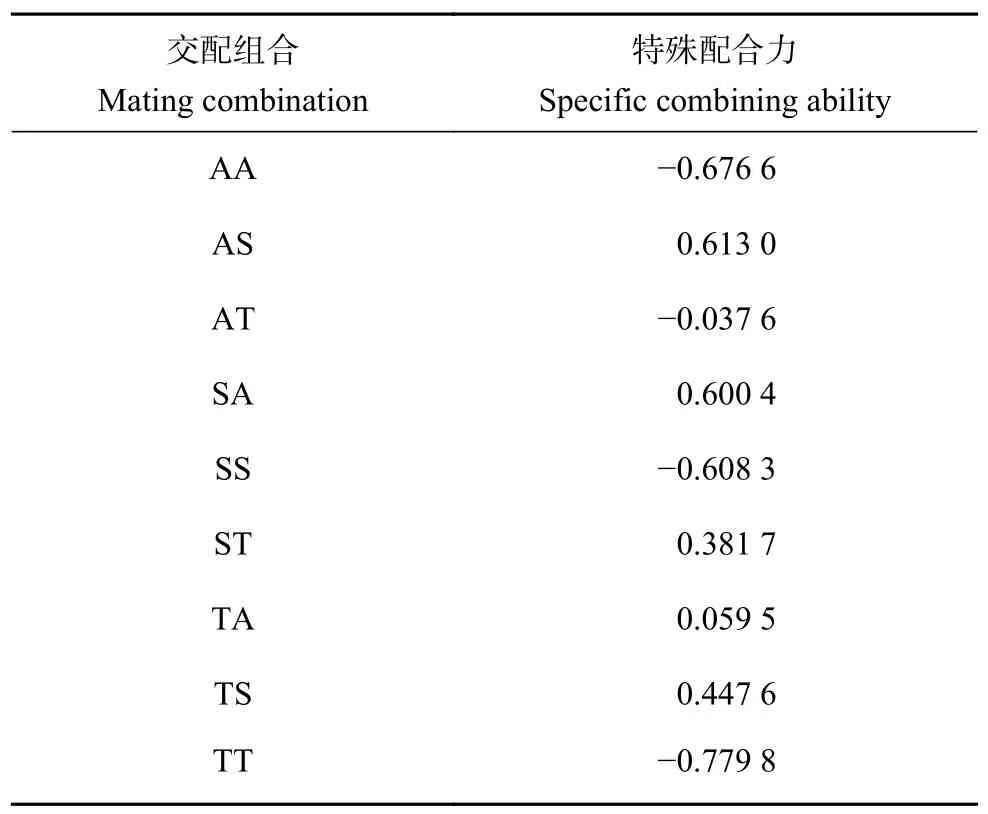
表4 9個(gè)交配組合體質(zhì)量的特殊配合力效應(yīng)值Table 4 Specific combining ability in harvest body mass of nine mating combinations
特殊配合力反映的是雙親雜交后通過互作表現(xiàn)出的基因非加性效應(yīng),即顯性效應(yīng)[27],是選取性狀優(yōu)良最佳雜交組合的重要依據(jù)。正常情況下,一般配合力高的種群雜交可以獲得好的雜交效果,其特殊配合力效應(yīng)值也高。通過本研究3 個(gè)群體的一般配合力和特殊配合力分析表明,2 個(gè)一般配合力高的“南海1 號(hào)”群體和非洲品系選育群體雜交組合的特殊配合力均為正值,且高于其他組合(表3、表4),其中以“南海1 號(hào)”群體為母本,非洲品系選育群體為父本的雜交組合 (AS) 的特殊配合力效應(yīng)值最高。這一結(jié)果表明該組合存在較強(qiáng)的非加性效應(yīng),在體質(zhì)量性狀方面雜交優(yōu)勢(shì)明顯。
斑節(jié)對(duì)蝦體質(zhì)量性狀配合力的方差組分見表5。體質(zhì)量性狀一般配合力方差和特殊配合力方差數(shù)值分別為0.452 2 和0.427 2,占表型方差比例分別為6.04%和5.70%。
2.3 3 個(gè)群體之間的雜種優(yōu)勢(shì)分析
已有研究發(fā)現(xiàn),無論兩個(gè)群體的親緣關(guān)系如何,雜交后代都會(huì)表現(xiàn)出不同程度的雜種優(yōu)勢(shì)[28]。斑節(jié)對(duì)蝦“南海1 號(hào)”、非洲品系以及泰國(guó)群體3 個(gè)群體之間的雜種優(yōu)勢(shì)分析表明,6 個(gè)雜交群體的體質(zhì)量均值為10.70 g,3 個(gè)自交群體的體質(zhì)量均值為9.56 g,體質(zhì)量總雜種優(yōu)勢(shì)為11.92%。各雜交群體體質(zhì)量的中親雜種優(yōu)勢(shì) (HM) 和超親雜種優(yōu)勢(shì) (HB) 見表6。所有雜交組合體質(zhì)量的中親雜種優(yōu)勢(shì) (HM) 和超親雜種優(yōu)勢(shì) (HB) 均為正值且存在正向雜種優(yōu)勢(shì),其中非洲品系群體 () ד南海1 號(hào)”群體 (♀) (AS) 的中親雜種優(yōu)勢(shì) (HM) 和超親雜種優(yōu)勢(shì) (HB) 數(shù)值最大,分別為13.80%和7.97%。

表5 體質(zhì)量性狀配合力的方差組分Table 5 Variance component of combining ability in harvest body mass
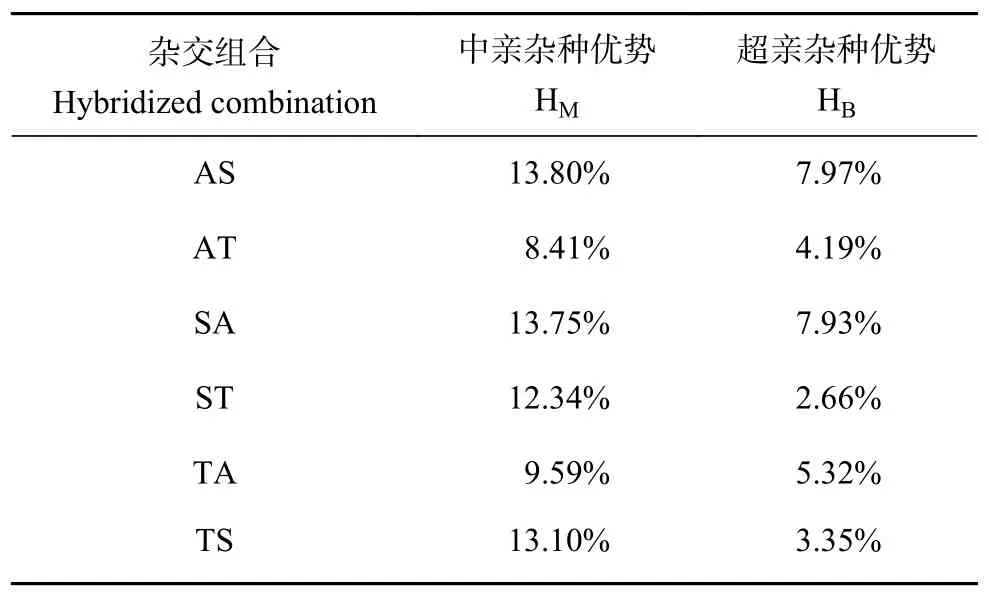
表6 6個(gè)雜交組合體質(zhì)量的中親雜種優(yōu)勢(shì)和超親雜種優(yōu)勢(shì)Table 6 Heterosis in harvest body mass of six hybridized combinations
目前,關(guān)于斑節(jié)對(duì)蝦不同群體之間的雜種優(yōu)勢(shì)分析不多。李永等[29]研究了三亞和泰國(guó)兩個(gè)地理群體的生長(zhǎng)性狀雜種優(yōu)勢(shì),發(fā)現(xiàn)雜交組合在體長(zhǎng)、體質(zhì)量等各個(gè)性狀上均表現(xiàn)出一定的雜種優(yōu)勢(shì)(2.13%~19.70%)。孫苗苗[30]研究了印度尼西亞和泰國(guó)兩個(gè)地理群體生長(zhǎng)性狀雜種優(yōu)勢(shì),發(fā)現(xiàn)生長(zhǎng)相關(guān)性狀存在一定的雜種優(yōu)勢(shì) (2.02%~8.50%)。本研究分析了“南海1 號(hào)”群體、非洲品系群體和泰國(guó)群體之間的雜種優(yōu)勢(shì),結(jié)果表明雜交子代在體質(zhì)量性狀上均表現(xiàn)出較大的雜種優(yōu)勢(shì) (表6),且明顯好于印度尼西亞和泰國(guó)群體之間體質(zhì)量性狀的雜種優(yōu)勢(shì)。
在雜交過程中,普遍存在正反交優(yōu)勢(shì)不同的現(xiàn)象[31],此現(xiàn)象可能與母性效應(yīng)、性別連鎖等有關(guān)[32-33],為雜交育種中采用特定的親本提供參考依據(jù)[32]。本研究結(jié)果表明,3 個(gè)群體之間正反交均表現(xiàn)出一定的雜交優(yōu)勢(shì)差異,但差異不明顯。在6 個(gè)雜交組合子代中,以“南海1 號(hào)”群體為母本要好于為父本的雜交優(yōu)勢(shì),其中“南海1 號(hào)”群體為母本,非洲品系為父本的雜種優(yōu)勢(shì)最好,因此,可以考慮加強(qiáng)該組合在體質(zhì)量方面的選育。
3 結(jié)論
本研究利用混合線性模型分析了“南海1 號(hào)”群體、非洲品系選育群體和泰國(guó)群體3 個(gè)斑節(jié)對(duì)蝦種質(zhì)群體之間的一般配合力和特殊配合力,并在此基礎(chǔ)上分析了3 個(gè)群體之間的中親和超親雜種優(yōu)勢(shì)。研究結(jié)果顯示“南海1 號(hào)”群體一般配合力最高,以“南海1 號(hào)”群體為母本,非洲品系選育群體為父本的雜交組合 (AS) 的特殊配合力效應(yīng)值最高,雜種優(yōu)勢(shì)也最為明顯。表明較非洲品系選育群體和泰國(guó)群體,“南海1 號(hào)”群體在質(zhì)量性狀方面存在較為豐富的加性遺傳變異;AS 組合存在較強(qiáng)的非加性效應(yīng),在體質(zhì)量性狀方面雜交優(yōu)勢(shì)明顯,可以考慮加強(qiáng)該組合在體質(zhì)量方面的選育。
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年12期)2022-02-14 07:08:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
