NaHS 引發提高裸燕麥種子活力的生理機制
2021-02-27 01:54:14劉建新劉瑞瑞賈海燕卜婷李娜
草業學報 2021年2期
劉建新,劉瑞瑞,賈海燕,卜婷,李娜
(甘肅省高校隴東生物資源保護與利用省級重點實驗室,隴東學院生命科學與技術學院,甘肅慶陽745000)
裸燕麥(Avena nuda)也稱莜麥,是起源于中國的一種禾本科燕麥屬一年生糧飼兼用作物,其栽培歷史已有兩千多年,在我國內蒙古、河北和山西等北方地區廣泛種植,播種面積僅次于小麥(Triticum aestivum)、水稻(Oryza sativa)和玉米(Zea mays)[1]。裸燕麥籽粒的蛋白質含量高達15.6%,脂肪含量達3.1%~11.6%,遠高于小麥、水稻、玉米和蕎麥(Fagopyrum esculentum);其富含膳食纖維、黃酮和皂苷等營養保健成分[2],具有降低血糖、血壓和血清膽固醇及改善胃腸功能的作用[3]。由于裸燕麥籽粒油脂含量高,且油脂中不飽和脂肪酸達80%以上[1],所以油脂酸敗容易導致種子發生劣變,從而降低種子活力。因此,研究提高裸燕麥種子活力的技術途徑對種質資源保存利用及提高播種質量具有重要意義。
種子引發(seed priming)是控制種子緩慢吸水并回干的一項播前種子處理技術[4],在1973 年由Heydecker等[5]提出,它通過控制種子緩慢吸水讓種子停留在吸脹階段,使種子處于預發芽的狀態,但胚根不會突破種皮。目前采用的種子引發技術主要有固體基質引發、生物引發、膜引發和液體引發等[6]。研究證明,引發不僅能提高種子萌發率[7],使出苗率提高和出苗整齊[8],而且還可增強種子和幼苗抵抗低溫[9]、水分脅迫[10]和鹽脅迫[11]等各種逆境的能力。目前應用較多的引發劑主要有FeSO4、PEG-6000、KNO3、GA3、多胺、茉莉酸甲酯、褪黑素和CaCl2等溶液[4,6?11],而以細胞氣體信使分子供體為引發劑進行種子引發的研究還少見報道。硫化氫(hydrogen sulfide,H2S)是繼NO 和CO 之后在植物體內發現的第3 種氣體信號分子,它參與植物生長發育和生理代謝過程調控[12]。已有研究表明,外源H2S 能夠促進小麥種子萌發[13];緩解干旱對水稻種子萌發的抑制[14];提高鹽脅迫下苜蓿(Medicago sativa)萌發種子抗氧化酶活性[15]和番茄(Lycopersicon esculentum)種子活力[16]。那么,以外源H2S 供體硫氫化鈉(NaHS)作為引發劑進行種子引發能否提高作物種子活力,以及相關的引發機制研究迄今未見報道。本研究以裸燕麥為材料,使用NaHS 進行種子引發,了解其對種子活力的影響,比較NaHS 引發種子與未引發種子在儲藏物質及抗氧化系統等方面的差異,揭示NaHS 種子引發的生理機理,為外源H2S 進行種子引發提供理論依據。
1 材料與方法
1.1 供試材料
試驗于2019 年4?6 月在甘肅省高校隴東生物資源保護與利用省級重點實驗室內進行。供試裸燕麥品種為‘定莜9 號’,種子由甘肅省定西市農業科學研究院提供。H2S 供體NaHS 購自Sigma?Aldrich 公司,其他化學試劑均為國產分析純。
1.2 試驗設計
1.2.1 NaHS 引發處理 挑選大小均一的裸燕麥健康種子約10 g 置于100 mL 燒杯中,分別加入0、50、100、200、400、800、1600 μmol·L?1NaHS 溶液,種子重量與溶液體積比為1︰3(g︰mL),在20 ℃避光引發18 h,蒸餾水洗凈殘余NaHS 后用吸水紙吸干表面水分,在室溫下回干至初始含水量7.3%時的重量后裝入磨口瓶中于冰箱中4 ℃保存備用。以未引發的裸燕麥種子(含水量7.3%)作對照(CK)一同進行發芽試驗,篩選NaHS 引發濃度。以篩選的800 μmol·L?1NaHS 溶液分別引發裸燕麥種子6、9、12、15、18 和21 h,蒸餾水沖洗后室溫回干至種子初始含水量(7.3%),然后進行發芽試驗,篩選NaHS 引發時間。試驗重復5 次。
用800 μmol·L?1NaHS 溶液20 ℃避光引發18 h,蒸餾水清洗后室溫回干至初始含水量(7.3%)的裸燕麥種子作為NaHS 引發種子;以蒸餾水(H2O)引發的種子作為NaHS 引發種子的陽性對照,排除H2O 的影響;以未引發的種子作為陰性對照(CK)。將NaHS 引發、H2O 引發和CK 種子4 ℃保存,立刻直接用于各項生理指標測定。每個處理5 次重復。
1.2.2 種子萌發指標測定 參照國家標準農作物種子檢驗規程GB/T3543.4-1995[17],將不同濃度NaHS 引發和NaHS 引發不同時間處理的種子分別均勻擺放在鋪有兩層濾紙的培養皿(直徑120 mm)中,每皿放置100 粒,各加5 mL 蒸餾水后在培養箱(LPGZ-250A,無錫萊浦儀器設置有限公司)中20 ℃、14 h/10 h(光/暗)培養。在種子置床后每12 h 觀察1 次,以胚根突破種皮2 mm 為發芽標準,記錄種子發芽數,每次觀察時補充適量蒸餾水,共觀察6 d。發芽結束后將幼苗在105 ℃殺青30 min 后65 ℃烘干至恒重稱量干重,計算發芽勢(germination poten?tial,GP)、發芽率(germination rate,GR)、發芽指數(germination index,GI)、活力指數(vigor index,VI)和幼苗單株干重(dry weight,DW)。GP(%)=前3 d 發芽種子數/供試種子數×100;GR(%)=發芽結束后的發芽種子數/供試種子數×100;GI=∑(Gt/Dt),Gt為第t 天的種子發芽數,Dt為相應的發芽天數;VI=GI×DW。
1.2.3 引發種子生理指標測定 采用亞甲基藍法測定H2S 含量[18];采用α-萘胺顯色法測定超氧陰離子(O2·ˉ)含量,采用硫代巴比妥酸法測定丙二醛(malondialdehyde,MDA)含量,采用電導率儀法測定質膜相對透性(rela?tive permeability of plasma membrane,RPPM)[19];按Sergiev 等[20]的方法測定H2O2含量;按陳建勛等[21]的方法測定超氧化物歧化酶(superoxide dismutase,SOD)、過氧化氫酶(catalase,CAT)、過氧化物酶(peroxidase,POD)和抗壞血酸過氧化物酶(ascorbate peroxidase,APX)活性。按楊穎麗等[22]的方法測定還原型抗壞血酸(reduced ascorbic acid,ASA)、脫氫抗壞血酸(dehydroascorbic acid,DHA)、還原型谷胱甘肽(reduced glutathione,GSH)和氧化型谷胱甘肽(oxidized glutathione,GSSG)含量。采用高俊鳳[19]的分光光度法、3,5-二硝基水楊酸顯色法、蒽酮比色法和考馬斯亮藍染色法分別測定細胞色素氧化酶(cytochrome oxidase,COX)和淀粉酶活性及淀粉、可溶性糖和可溶性蛋白質含量。
1.3 數據統計與分析
用SPSS 20.0 對數據進行單因素方差分析,對百分率數據反正弦轉換后進行方差分析,采用Duncan 法多重比較(P<0.05),數據結果以平均值±標準差表示。
2 結果與分析
2.1 不同濃度NaHS 引發對裸燕麥種子活力的影響
從表1 可見,與不引發的CK 相比,不同濃度NaHS 引發對裸燕麥種子的發芽指標影響不同。其中800 μmol·L?1NaHS 引發顯著提高了GP、GR、GI 和VI,但對DW 的影響不大。另外,除100 μmol·L?1NaHS 引發顯著提高了GR、200 μmol·L?1NaHS 引發顯著提高了GI 外,其他各濃度NaHS 引發的GP、GR、GI、VI 和DW 均與CK 的差異不顯著。因此,選用800 μmol·L?1作為后續試驗NaHS 的引發濃度。
2.2 NaHS 不同引發時間對裸燕麥種子活力的影響
從800 μmol·L?1NaHS 引發不同時間對裸燕麥種子發芽指標的影響可以看出(表2),除引發21 h 的GP 顯著高于12 h、引發12 h 的GR 顯著低于引發15、18 和21 h 外,其他各引發時間的GP 和GR 無顯著差異;除引發18 和21 h 的GI、VI 顯著高于引發6、9、12 和15 h 外,其他各引發時間的GI 和VI 差異不顯著;不同引發時間的DW 差異均不顯著。由于引發18 h 的GI 和VI 顯著高于引發6 ~15 h 的GI 和VI,且GP、GR 和DW 相對較大,并與引發21 h 的GP、GR、GI、VI 和DW 無顯著差異,所以選用引發18 h 作為NaHS 引發時間。

表1 不同濃度NaHS 引發對裸燕麥種子發芽指標的影響Table 1 Effect of seed priming with different concentrations of NaHS on germination indexes of naked oat

表2 用800 μmol·L-1 NaHS 引發不同時間對裸燕麥種子發芽指標的影響Table 2 Effect of 800 μmol·L-1 NaHS priming times on germination indexes of naked oat seeds
2.3 NaHS 引發對裸燕麥種子貯藏物質含量的影響
表3 表明,與CK 相比,NaHS 和H2O 引發均未顯著改變裸燕麥種子淀粉和可溶性蛋白質含量;H2O 引發顯著降低了種子可溶性糖含量,NaHS 引發的種子可溶性糖含量與CK 和H2O 引發無顯著差異。
2.4 NaHS 引發對裸燕麥種子細胞色素氧化酶和淀粉酶活性的影響
細胞色素氧化酶(COX)是電子傳遞鏈末端酶,淀粉酶是催化淀粉水解的關鍵酶。表4 表明,NaHS 引發顯著提高了裸燕麥種子的COX 活性,分別比CK 和H2O 引發提高了103.3%和19.4%,H2O 引發的COX活性比CK 提高了70.3%,且差異顯著。與CK 相比,NaHS 和H2O 引發均顯著降低了α-淀粉酶、β-淀粉酶和(α+β)淀粉酶活性,α-淀粉酶活性分別降低了53.6%和43.9%,β-淀粉酶活性分別降低了39.8%和16.4%,(α+β)淀粉酶活性分別降低了40.1%和17.1%。NaHS 引發與H2O 引發相比,α-淀粉酶活性無顯著差異,β-淀粉酶和(α+β)淀粉酶活性分別下降了27.9%和27.7%,且差異顯著。
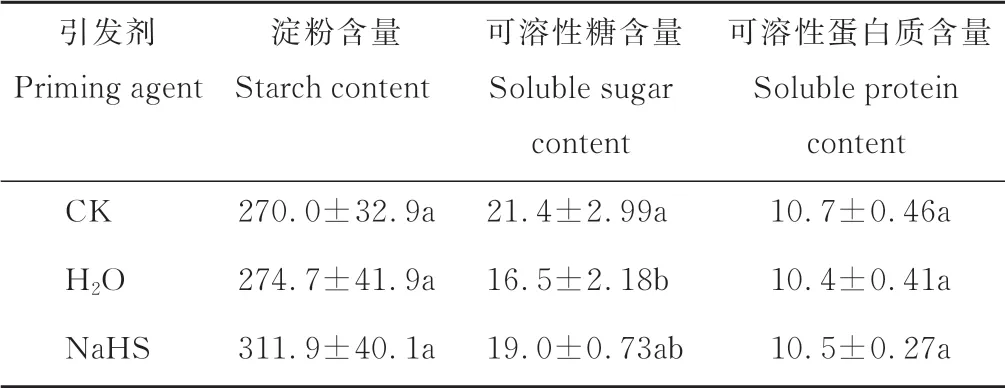
表3 不同引發劑對裸燕麥種子淀粉、可溶性糖和可溶性蛋白質含量的影響Table 3 Effect of seed priming with different agents on the contents of starch,soluble sugar and soluble protein of naked oat seeds(mg·g-1)

表4 不同引發劑對裸燕麥種子細胞色素氧化酶和淀粉酶活性的影響Table 4 Effect of seed priming with different agents on the activities of cytochrome oxidase and amylase of naked oat seeds
2.5 NaHS 引發對裸燕麥種子H2S 含量、活性氧產生和膜脂過氧化的影響
由表5 可見,NaHS 引發顯著提高了裸燕麥種子中H2S 的含量,比CK 提高了113.5%,比H2O 引發提高了79.7%,而H2O 引發種子的H2S 含量與CK 差異不顯著。NaHS 引發顯著降低了O2·ˉ、H2O2、MDA 含量和RPPM,分別比CK 下降了34.7%、36.1%、37.6%和11.9%,比H2O 引發分別下降了11.7%、22.6%、29.0%和7.3%;與CK 相比,H2O 引發顯著降低了O2·ˉ含量和RPPM,分別下降了26.0%和4.6%,而對H2O2和MDA 含量無顯著影響。

表5 不同引發劑對裸燕麥種子H2S、O2·ˉ、H2O2、MDA 含量和質膜相對透性的影響Table 5 Effect of seed priming with different agents on contents of H2S,O2·ˉ,H2O2,MDA and RPPM of naked oat seeds
2.6 NaHS 引發對裸燕麥種子抗氧化酶活性的影響
由表6 可知,NaHS 引發顯著提高了裸燕麥種子SOD、CAT 和POD 活性,分別比CK 提高了6.1%、112.0%和120.5%,比H2O 引發分別提高了6.2%、85.8%和63.7%,而其APX 活性與CK 和H2O 引發的差異不顯著。與CK 相比,H2O 引發對SOD 和CAT 活性無顯著影響,但POD 和APX 活性分別顯著提高了34.7%和49.1%。

表6 不同引發劑對裸燕麥種子SOD、CAT、POD 和APX 活性的影響Table 6 Effect of seed priming with different agents on activities of SOD,CAT,POD and APX of naked oat seeds
2.7 NaHS 引發對裸燕麥種子抗氧化物質含量的影響
由表7 可知,NaHS 引發的裸燕麥種子中ASA、DHA 含量和ASA/DHA 均與CK 和H2O 引發的無顯著差異,卻顯著降低了GSH 含量和GSH/GSSG,分別比CK 下降了29.2%和38.1%,GSSG 含量卻比CK 顯著提高了14.4%。NaHS 引發的種子GSH、GSSG 含量和GSH/GSSG 與H2O 引發的差異不顯著。

表7 不同引發劑對裸燕麥種子抗氧化物質含量的影響Table 7 Effect of seed priming with different agents on the antioxidant contents of naked oat seeds
3 討論
引發是作物播前種子預處理的一項重要措施。研究證明,引發能夠增強種子活力和提高幼苗抗逆性[7?11]。研究者通常用發芽勢(GP)、發芽率(GR)、發芽指數(GI)和活力指數(VI)等發芽指標反映引發種子的活力狀況[11]。GP 反映種子發芽速率;GR 反映在一定時間內總的種子發芽百分率;GI 既能反映發芽速率,又能反映發芽的整齊程度;VI 是發芽速率和生長量的綜合反映,是衡量種子活力更好的指標[23]。現有的種子引發研究多用低滲透勢高分子物質或小分子滲透物質作為引發劑[4,6?11],以外源信號分子供體作引發劑進行種子引發的研究尚鮮見報道。本研究發現,用800 μmol·L?1外源H2S 供體NaHS 作為引發劑引發18 h 顯著提高了裸燕麥種子的GP、GR、GI 和VI,但對幼苗生長量的影響不大(表1 和表2)。這與Zhang 等[13]用NaHS 預處理12 h 可促進小麥種子萌發的結果類似。說明用氣體信號H2S 供體引發可提高裸燕麥的種子活力。
關于種子引發機理的研究大多從引發種子萌發苗生理生化的變化進行探討,通過引發種子內部變化揭示引發生理機制的研究尚鮮見報道[4]。種子引發通常是種子在低水勢溶液中緩慢吸水后再回干至初始狀態的過程,這個過程中可能會使種子內部的相容性溶質(compatible solutes)如脯氨酸、甜菜堿和可溶性糖等含量發生改變。阮松林等[24]研究表明,用脯氨酸和NaCl 引發可提高水稻種子脯氨酸和蔗糖含量,而降低果糖和可溶性糖含量。牛曉雪等[4]發現,FeSO4引發能夠降低秦艽(Gentiana macrophylla)種子中蔗糖和可溶性蛋白質含量,對淀粉、葡萄糖和果糖含量無影響。本研究中,用NaHS 引發并未引起裸燕麥種子中淀粉、可溶性糖和可溶性蛋白質含量的顯著變化(表3)。這與王彥榮等[7]用PEG 引發紫花苜蓿和沙打旺(Astragalus adsurgens)種子在萌發吸水初始階段可溶性糖和脯氨酸含量無顯著變化的結果類似。說明NaHS 引發并不是通過提前啟動物質代謝來增強裸燕麥的種子活力。進一步研究發現,NaHS 引發顯著降低裸燕麥種子中α-淀粉酶、β-淀粉酶和總淀粉酶活性(表4)。這與前人NaHS 處理可提高小麥種子胚乳β-淀粉酶活性,而對α-淀粉酶合成和活性沒有影響的結果[13]有區別。這可能與不同植物種類響應機制存在差異有關。淀粉酶是一類催化種子貯藏淀粉水解成小分子糖的關鍵酶。有研究表明,H2S 參與植物體內關鍵半胱氨酸(cysteine,Cys)殘基的巰基化修飾,從而調節蛋白質的活性[18]。本研究中,NaHS 引發顯著提高了裸燕麥種子中H2S 的含量(表5)。NaHS 引發抑制裸燕麥種子淀粉酶活性的原因可能與H2S 參與Cys 殘基的巰基化修飾有關。因為引發是種子緩慢吸水后仍讓種子回干至原來狀態的過程,并未讓種子處于萌發階段。這時H2S 可能通過修飾抑制淀粉酶的活性,不讓種子啟動萌發。本研究中NaHS 引發未引起裸燕麥種子貯藏物質含量變化的結果與此相互佐證,進一步證明NaHS 引發并非通過貯藏物質動員來提高種子活力。但NaHS 引發抑制淀粉酶活性的具體機制有待進一步揭示。線粒體是細胞物質和能量代謝的場所,其內膜上的COX 活性反應細胞有氧代謝的水平[25]。本研究中,NaHS 引發顯著提高了裸燕麥種子中的COX 活性(表4)。這與用FeSO4引發提高秦艽種子COX 活性的結果相一致[4]。由此認為,NaHS 引發增強線粒體呼吸代謝水平可能是提高種子活力的生理調節機制之一。若線粒體有氧呼吸水平降低,便會產生呼吸鏈電子泄漏,導致活性氧積累造成線粒體結構破壞[25]。本研究結果顯示,NaHS 引發能顯著降低裸燕麥種子中2 種主要活性氧(O2·ˉ、H2O2)的含量,也使細胞膜脂過氧化產物MDA 含量和RPPM 明顯下降(表5),說明NaHS 引發可通過降低活性氧水平維持細胞結構和功能的完整性。這與王彥榮等[7]和楊小環等[26]的引發可促進種子細胞膜修復,降低內容物外滲的研究結果一致。
引發本身對種子就是一種脅迫,能夠誘導細胞防御體系的建立[4]。植物體內的抗氧化防御系統在維持活性氧代謝平衡、防止膜脂氧化損傷方面發揮著重要作用[14]。抗氧化系統包括酶系統和抗氧化物質,其中SOD 催化O2·ˉ發生歧化反應產生H2O2,H2O2進一步被CAT、POD 和APX 協同轉化成H2O[16];ASA 和GSH 則通過ASA?GSH 循環參與H2O2的清除[22]。本研究結果發現,NaHS 引發顯著提高裸燕麥種子中的SOD、CAT 和POD 活性,降低GSH 含量和GSH/GSSG,而對APX 活性及ASA、DHA 含量和ASA/DHA 的影響不大(表6 和表7)。由此認為,NaHS 引發主要通過提高SOD、CAT 和POD 等抗氧化酶活性來維持裸燕麥種子較低水平的活性氧,從而降低對膜脂的氧化傷害程度,提高種子活力。這與外源NaHS 處理提高抗氧化酶活性緩解鹽脅迫對番茄種子萌發抑制[16]的結果基本一致。本研究中,NaHS 引發引起裸燕麥種子GSH/GSSG 降低的原因可能是引發促進了種子中已有GSH 參與了活性氧清除而被氧化成GSSG,進而通過ASA?GSH 循環維持了ASA/DHA 的穩定(表7)。由于ASA/DHA 相對穩定,NaHS 引發可能對APX 活性也表現為無顯著的影響(表6),但NaHS 引發激活SOD、CAT 和POD 活性的機制以及對ASA?GSH 循環相關酶系的調節作用尚待進一步探究。
4 結論
用800 μmol·L?1NaHS 引發18 h 可以提高裸燕麥的種子活力。NaHS 引發提高裸燕麥種子活力的可能生理機制:一是引發提高種子內H2S 含量,進而誘導SOD、CAT 和POD 等抗氧化酶活性提高,通過GSH 氧化成GSSG 維持ASA/DHA 和APX 活性的相對穩定,從而有效清除O2·ˉ和H2O2等活性氧,使膜脂過氧化程度保持在較低水平;二是通過激活線粒體COX 活性,增強細胞有氧呼吸代謝水平,從而提高種子活力。NaHS 引發并非通過激活淀粉酶活性提前啟動淀粉等貯藏物質轉化而提高裸燕麥種子活力。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
