黃河源人工草地土壤微生物多樣性對建植年限的響應
2021-02-27 01:53:56孫華方李希來金立群李成一張靜
草業學報 2021年2期
孫華方,李希來,2*,金立群,李成一,張靜,2
(1.青海大學農牧學院,青海 西寧810016;2.青海-廣東自然資源監測與評價聯合實驗室,青海 西寧810016)
土壤微生物是土壤生態系統的重要組成部分,對土壤中有機化合物的降解、礦化和腐殖化有顯著作用[1?2],不僅在全球生物地球化學循環中占據中心位置[3],其群落結構和多樣性對土壤功能和草地生產力也至關重要。研究發現,不同的微生物通過分泌不同胞外酶從而在土壤養分循環中發揮著不同的功能。如真菌能夠通過釋放磷轉化酶增加土壤磷含量,而細菌中含有多種固氮菌種對土壤中氮積累有顯著作用,所以真菌和細菌能夠共同促進土壤氮磷平衡[4?5]。有學者指出,微生物群落在沙漠地區生物化學循環過程中十分重要,尤其是在碳氮循環中。也有學者認為某些微生物會與植被形成共生體系,如固氮菌與豆科植物形成根瘤菌,共同促進彼此生長[2,6?7],在高寒地區,陳懂懂等[8]對一年不同混播草地土壤微生物變化研究發現,種植禾本科植物有利于微生物多樣性恢復,但在人工草地長期演替過程中各演替階段表層土壤微生物的變化特征及生態作用還不清楚。
此外,微生物結構能夠表征草地土壤狀況[9]。當微生物結構失調時草地狀態可能發生變化。草地退化不利于土壤團聚體形成,會加劇土壤養分流失,導致適應性差的菌種消失。在三江源地區,Che 等[10]發現,退化會降低高寒草甸微生物多樣性和活性,同時草地退化也會改變某些微生物代謝途徑,分泌有毒物質對植物生長產生不利影響[11?13]。黃河源高寒地區人工草地由于不合理的利用,造成的退化已經是普遍現象。目前對該地區人工草地監測大多集中在植被、土壤理化性質和微生物數量變化上[14?16],而對環境變化更敏感、具有指示性的微生物多樣性變化特征知之甚少,所以對人工草地演替過程中土壤微生物結構變化的研究在監測人工草地恢復中具有很重要的意義。
有大量學者已經探討了土壤理化性質如土壤養分、增溫等對微生物的影響,也有少量學者針對不同碳沉降條件下微生物的變化進行研究,但大多都集中在高寒草甸和恢復年限短的人工草地上,同時僅利用常規分析法對三大類微生物(真菌、細菌、放線菌)數量變化進行探究[17?20],對于高寒地區恢復年限長達18 年的人工草地中的微生物多樣性研究較少。此外,人工草地演替過程中植被特征變化較土壤更加敏感[21],其是否會對微生物產生更大的影響還不清楚,所以本研究著重討論植被變化對微生物的影響,能夠及時準確預測微生物在人工草地演替過程中的變化特征。基于此,本研究選取黃河源區不同恢復時間人工草地,調查表層土壤微生物群落結構特點以及分析微生物與植被因子的相關性,了解人工草地表層土壤微生物群落組成對植被和建植年限的響應,為黃河源區人工草地的管理提供科學依據。
1 材料與方法
1.1 研究區域概況
研究樣地位于瑪沁縣,地理位置98°45′?100°56′E,33°43′?35°16′N。地處青海省東南部,果洛藏族自治州的東北部阿尼瑪卿山區,屬典型的高原山地地貌,平均海拔高度4100~4500 m。具有青藏高原寒冷氣候特點,冬季長而干寒,夏季短而溫濕。年平均氣溫為?3.9~3.8 ℃,極端最低氣溫?31.4 ℃;年降水量492.7 mm,是青海省降水最多的地區之一,無絕對無霜期。
選擇瑪沁縣大武灘上不同種植年限人工草地(18,14,11,4,1 年)作為調查對象,人工草地牧草混播品種有垂穗披堿草(Elymus nutans)、冷地早熟禾(Poa crymophila)、中華羊茅(Festuca sinensis),混播比例為2∶1∶1。所用草種均由當地草籽繁殖場生產,播量為45 kg·hm?2,施肥量為45 kg·hm?2(磷酸二銨復合肥),使用C-型肉毒素控制高原鼠兔種群數量[22?23]。人工草地建植之前屬于高寒沼澤濕地外周典型“黑土灘”退化草地,優勢種莎草科植物被毒雜草取代,原生植被不足10%,裸露地大面積存在。人工草地生長季完全禁牧,只冬季放牧利用(每年12 月至次年4 月)。試驗樣地地理位置見圖1,試驗地概況詳見文獻[24]。
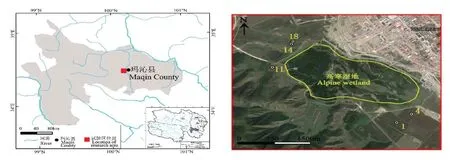
圖1 試驗樣地地理位置[24]Fig.1 Geographical location at study area
1.2 研究方法
1.2.1 土壤樣品采集 2018 年8 月在青海省瑪沁縣大武灘選擇5 個不同年限人工草地,調查植被和土壤微生物分布特征,樣地選擇和植被調查方法詳見孫華方等[24]的文獻報道。采用多點土壤混合法獲取土壤樣品,具體方法如下:植被特征調查后,使用醫用酒精浸泡后的脫脂棉擦拭鐵勺達到消毒目的,使用消毒后的鐵勺在樣方內隨機挖取表層(0~5 cm)土壤樣品裝于無菌袋中,重復5 次,將同一樣線土壤混合為1 個樣品,除11 年重復3 次外,其余各建植年限人工草地均重復4 次。共獲得19 個土壤樣品(11 年3 個樣品,其他4 個年限各4 個樣品)。將獲得的土壤樣品暫時保存在4 ℃便攜式冰箱內(BC-92BV,美國),再迅速運回實驗室于?80 ℃冰箱保存。
1.2.2 微生物測定方法 DNA 提取:使用Power Soil DNA Isolation Kit(MoBio Laboratories,Carlsbad,CA)試劑盒提取基因組DNA,并使用Nanodrop 檢驗DNA 質量和濃度。PCR 擴增:使用KAPA 2G Robust Hot Start Ready Mix 及帶有錯位堿基的融合引物進行PCR 擴增(原核微生物擴增區域為16s V3?V4 區,引物為336F,806R;真核微生物擴增區域為ITS1 區,引物為ITS1-F,ITS2),PCR 擴增總體系為25 μL。使用1%瓊脂糖凝膠電泳檢測擴增目的條帶大小(170 V,30 min),并使用磁珠法進行自動化純化。Miseq 文庫構建:按照Pooling比例,對沒有錯位堿基的融合引物PCR 產物取一定體積Pooling 成一個上機文庫,并使用2%瓊脂糖凝膠對文庫片段進行篩選。篩選后的文庫片段經Qubit 檢測定量,取一定量文庫加入10 μL End repair & Add A 進行末端修復加A 尾,再加入33.5 μL Adaptor Ligation Mix 進行加測序接頭的連接,并對文庫進行純化回收。然后加入接頭引物、酶及Mix 進行PCR 富集完成文庫構建。最后,對文庫進行PCR 產物純化。上機測序:文庫在Illumina Miseq 平臺上機測序。以上過程均委托梅塔生物公司(甘肅)完成。
1.3 數據處理與分析
本研究數據利用Excel 進行整理。采用Qiime 軟件,將相似度達97%以上的序列歸為一個操作分類單元即OTU;聚類所有序列(Ucl 法),參照RDP(ribosomal database project)數據庫,采用貝葉斯算法注釋每個分類中的OTU 代表序列,得到每個OTU 的分類學信息。對OTU 數據進行整理,在微生物分類為門、屬的水平上對樣品中真核、原核微生物豐度進行統計,利用Excel 繪柱狀圖。同時,利用SPSS 19.0 對植被特征與微生物OTUs 數量進行單因素方差分析;對植被和微生物特征(原核、真核微生物OTUs 數量、微生物多樣性)進行皮爾遜(Pearson)相關性分析,使用雙尾檢測,P<0.05 時顯著相關,P<0.01 時極顯著相關。
微生物多樣性計算公式:

式中:Pi指第i種占總數的比例;H′為多樣性指數;J′為均勻度指數;S為群落中微生物的種數;D為生態優勢度指數;Schao1為估計的OTU 數;Sobs為觀測到的OTU 數;n1為只有一條序列的OTU 數目;n2為只有兩條序列的OTU數目。
2 結果與分析
2.1 不同建植年限人工草地土壤微生物數量分布特征
通過分析不同建植年限人工草地表層(0~5 cm)土壤微生物的OTUs 數量發現,原核微生物OTUs 數量變化范圍是15696~21019 個,總體趨勢為隨著建植年限的增長,原核微生物OTUs 數量增多。真核微生物變化表現出相似的規律。建植1 和4 年草地中原核、真核微生物OTUs 數量均顯著低于其他年份人工草地(P<0.05)。總體而言,隨著人工草地建植年限增加,土壤微生物OTUs 數量逐漸恢復。對比真核和原核微生物發現,各相同年限的人工草地土壤中真核微生物OTUs 數量均高于原核微生物,其中建植年限短(1,4,11 年)的人工草地之間差異顯著(P<0.05)。隨著人工草地的演替,原核和真核OTUs 數量差異減小。這一結果說明微生物群落結構隨著人工草地建植時間的增加而逐漸優化(圖2)。
2.2 人工草地恢復過程中土壤微生物群落結構特征
通過對不同恢復年限人工草地表層土壤微生物豐度分析發現:針對原核微生物(圖3I),各年限人工草地分別具有門22 個(18 年)、25 個(14 年)、23 個(11 年)、20 個(4 年)和21 個(1 年)。3 大優勢菌有變形菌門(Proteobacte?ria)、放線菌門(Actinobacteria)和酸桿菌門(Acidobacteria),占總數量的80%以上。各年限變形菌門和放線菌門微生物相對豐度均在25%以上,其中放線菌門微生物隨著建植年限增長相對豐度變大,但變形菌門與年限相關性差。酸桿菌門微生物相對豐度均在20%以上。其他菌門如芽單胞菌門(Gemmatimonadetes)、擬桿菌門(Bacte?roidetes)微生物等相對豐度相對較低,在3%~7%。相對豐度<1%的菌門歸為其他(other)類型,其豐度約為2%左右。各人工草地原核微生物優勢菌門具有高度相似性,可能是由于這些優勢菌適應性強,能廣泛生存在不同土壤中。但由于環境的差異性,不同人工草地樣地各優勢菌門組成比例不同,但隨著建植年限的增加,各有益菌門不斷增加,故人工草地建植過程中,土壤原核微生物正在逐漸恢復。
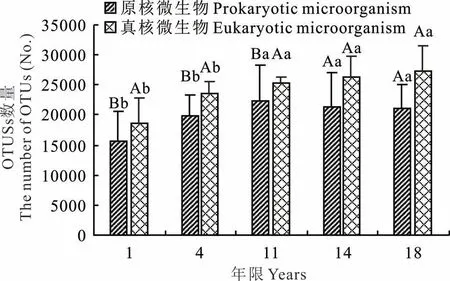
圖2 不同恢復年限人工草地土壤微生物OTUs 數量Fig.2 The number of soil microbial OTUs within different years in artificial grassland
就真核微生物而言(圖3II),不同年限人工草地微生物門的種類少于原核微生物,其中優勢門均為子囊菌門(Ascomycota)和擔子菌門(Basidiomycota)。不同年限人工草地子囊菌門微生物相對豐度均在64%以上,其中11年人工草地最高,為84.99%。針對擔子菌門,相對豐度變化范圍為9.07%~21.65%,總體變化趨勢是隨建植年限增加而增加。子囊菌門作為人工草地土壤微生物中的絕對優勢菌,該門中有很多有害微生物如赤霉屬(Gib?berella),對植被及其他微生物產生不利影響。隨著恢復年限的增加,該菌門相對豐度逐漸減少,有害微生物減少,有利于土壤健康的發展。綜上所述,有益菌隨著人工草地建植時間的增長逐漸增加,而有害菌相對豐度逐漸減少,因此土壤微生物群落隨人工草地的恢復演替也在逐漸恢復。
通過對不同年限原核微生物屬水平的研究發現(圖4I):不同年限人工草地原核微生物屬的數量相差不大。各人工草地土壤原核微生物相對豐度小于1%的屬最多,其中11 年人工草地最多,為43.17%,其他年限人工草地在34%左右。說明在草地恢復過程中,更多類型的原核微生物在逐漸恢復。各年限原核優勢屬均為芽單胞菌屬(Gemmatimonas)、Gp4 屬、紅桿菌屬(Solirubrobacter)。就Gp4 屬而言,隨建植年限增加其相對豐度也在增加,在18 年人工草地相對豐度最高,為9.57%,其他年限差異不大,為7%~8%。但是芽單胞菌屬變化趨勢相反。紅桿菌屬隨著建植年限增加其相對豐度呈增加趨勢,18 年最高,為5.83%。
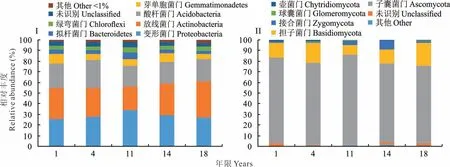
圖3 不同建植年限人工草地土壤微生物在門水平上的相對豐度Fig.3 Relative abundance of soil microorganisms at phylum level within different years in artificial grassland

圖4 不同建植年限人工草地土壤微生物在屬水平上的相對豐度Fig.4 Relative abundance of soil microorganisms at generic level within different years in artificial grassland
通過對不同年限真核微生物優勢屬研究發現(圖4II),真核微生物未鑒定出的屬最多,不同年限人工草地中均在21%以上,其中14 年人工草地相對豐度最高,為30.7%。所以,人工草地演替過程中土壤真核微生物結構和功能還需更深入的研究。雖然不同建植年限人工草地中真核微生物主要優勢屬相同,但各屬相對豐度如孢霉屬(Mortierella)等相差較大。
由表1 可知,不同建植年限人工草地原核、真核微生物多樣性具有一定差異,對比不同年限原核、真核微生物物種數發現:原核微生物物種數隨建植年限增加稍有下降。其中1 年人工草地中原核微生物物種數最多,為3100個,11 年人工草地中原核微生物物種數最少,為2047 個。11 年人工草地中原核微生物物種數分別與其他年限差異顯著(P<0.05)。其他年限之間原核微生物物種數沒有較大差異。真核微生物隨建植年限增加表現出相同趨勢,變化范圍為462~541 個,各年限人工草地之間真核微生物的物種數差異不顯著。通過對比相同年限人工草地中原核、真核微生物的物種數發現,各年限人工草地中原核微生物物種個數均顯著大于真核微生物物種個數(約5 倍,P<0.05)。
分析不同年限人工草地微生物多樣性指數發現,1 年原核微生物Shannon?Wiener 指數最大,為10.56,11 年最小,為9.65,彼此之間差異顯著(P<0.05),其他年限人工草地之間差異不顯著(表1)。就真核微生物的Shan?non 指數而言,14 年人工草地中最高,為6.45。建植14 年后Shannon?Wiener 指數發生下降趨勢,18 年人工草地Shannon 指數最小,為5.77。各年限人工草地中原核微生物的Shannon?Wiener 指數均顯著高于真核微生物(約2 倍,P<0.05)。
從不同人工草地土壤微生物Simpson 指數變化看,不同年限人工草地微生物Simpson 指數變化不大,均在1左右,但各年限人工草地原核、真核微生物的Chao1 指數差異較大,各年限原核微生物Chao1 指數均顯著大于真核微生物(約6~8 倍,P<0.05),其中1 年原核、真核微生物Chao1 指數相差最大,原核微生物Chao1 指數大約是真核微生物的7.7 倍(表1)。綜上所述,原核微生物物種數、多樣性指數(除Simpson 指數)均顯著大于真核微生物(P<0.05),表明人工草地中原核微生物多樣性恢復速度較快。

表1 不同恢復年限人工草地微生物多樣性特征Table 1 Microbial diversity within different years in artificial grassland
2.3 植被與微生物特征相關性分析
由表2 可知原核微生物OTUs 數量與大多人工草地植被特征(植被數據參考文獻[24],下同)密切相關,如與雜類草蓋度、植被多樣性指數(Simpson 指數、Shannon?Wiener 指數、Pielou 均勻度指數)極顯著相關(相關系數:0.626,0.612,0.715,0.631,P<0.01),與植被總蓋度、物種數顯著正相關(0.548,0.575,P<0.05)。而真核微生物OTUs 數量主要與生物土壤結皮(biological soil crusts,BSCs)蓋度、總蓋度(BSCs 數據參考文獻[24],下同)極顯著正相關(0.732,0.621,P<0.01),與禾本科蓋度顯著正相關(0.546,P<0.05),與其他植被特征如莎草科蓋度相關性較差。
選取優勢原核、真核微生物屬,分析人工草地植被特征與其相關性結果發現(表3),人工草地植被蓋度特征與原核微生物屬的相對豐度之間相關性不顯著(P>0.05),但植被總蓋度、BSCs 蓋度與主要真核微生物優勢屬Knufia顯著相關(0.503,0.494,P<0.05)。通過分析植物多樣性指數(Simpson 指數、Shannon?Wiener 指數、Pielou 均勻度指數)與原核、真核微生物優勢屬的相關性發現:真核微生物Davidiella屬與植物多樣性指數相關性很強,其中與Simpson 指數、Pielou 均勻度指數極顯著負相關(?0.677,?0.582,P<0.01),與Shannon?Wiener指數顯著負相關(?0.525,P<0.05),而原核微生物優勢屬與植被多樣性相關性較差。綜上所述,人工草地恢復演替過程中,植被可能對真核微生物的恢復作用較顯著。這主要是由于真核微生物對禾本科人工草地中高C/N的凋落物更敏感[25],故表現出顯著的相關性。

表2 不同年限人工草地植被特征與微生物OTUs 數量特征相關性分析Table 2 Correlation analysis between vegetation characteristics and OTUs

表3 不同年限人工草地植被特征與主要微生物相對豐度相關性分析Table 3 Correlation analysis between vegetation characteristics and main microorganisms
由表4 可知:莎草科植物蓋度與真核微生物Simpson 指數呈顯著負相關關系(?0.496,P<0.05);植被Simp?son 指數分別與真核微生物物種數和Chao1 指數顯著正相關(0.522,0.534;P<0.05),而植被Shannon?Wiener指數與真核微生物Chao1 指數顯著正相關(0.496,P<0.05)。人工草地植被特征與土壤原核微生物多樣性指數沒有較好的相關關系。
3 討論
3.1 植被對人工草地土壤微生物OTUs 數量的影響
微生物OTUs 數量是研究土壤微生物的最基本也很重要的一個指標。OTUs 是人為設定的分類單元,通常不同OTUs 代表不同微生物物種,OTUs 數量越多,微生物種類越多。在本研究中,隨著人工草地建植年限的增長,微生物OTUs 數量逐漸增多,與舒維花等[26]的結果相似。很多因子均能影響草地微生物的分布。夏北成[27]研究發現,植被能夠增加微生物多樣性,且微生物多樣性隨著人工草地建植年限增加而變大,本研究也得出相似結論。相關性分析表明植被多樣性與原核微生物多樣性關系緊密,該結果與蒲琴等[28]和薛凱等[29]的研究結果一致。可能是因為在人工草地演替過程中,維管束植物的恢復為微生物生長提供了良好環境,促進其多樣性增加[30]。此外,在草地生態系統中植被凋落物和根系分泌物是土壤微生物的主要碳源[31]。孫華方等[24]提出,隨著人工草地建植年限增加,植被多樣性也增加。多樣化的植物群落凋落物、根系分泌物直接或間接促進微生物群落多樣性發展[20,32?35],所以植被多樣性高的人工草地土壤微生物多樣性也高。BSCs 是人工草地恢復過程中除維管束植物外十分重要的伴生種,在降低維管束植物間裸地面積與退化土壤生態系統修復和土壤穩定性維持上作用顯著[36?39],土壤微生物作為BSCs 重要組成部分,在土壤微生物恢復中BSCs 具有重要意義。相關性分析得出BSCs蓋度與真核微生物OTUs 數量顯著正相關,所以BSCs 的恢復可能對土壤真核微生物的恢復有顯著作用,該結果與李發祥等[40]的結論一致。植被影響微生物的同時,土壤微生物對地上植被的生長也有強烈的反應。例如,土壤微生物能夠加速養分回歸土壤以及降解土壤中穩定性高的有機物[41?42],為植被提供更多可利用的速效養分從而加速植物生長。此外,某些微生物會與植被根系形成菌根,達到共同生長[7]。所以在草地生態系統中,地上植被和土壤微生物群落相互作用,共同影響高寒人工草地的群落演替和土壤恢復過程和特性。

表4 不同年限人工草地植被特征與微生物多樣性相關性分析Table 4 Correlation analysis between vegetation characteristics and microbial diversity
隨著人工草地建植年限增加,微生物OTUs 數量也增加,但是人工草地土壤微生物群落結構依然不平衡。通過對比人工草地表層土壤中原核、真核微生物OTUs 數量發現,建植18 年人工草地土壤依然處于非健康狀態,該結果(細菌/真菌<1)與彭岳林等[43]的研究結果不同,可能是因為人工草地狀況較差引起真核、原核微生物結構失衡。研究表明,微生物分布的異質性很大程度取決于周圍環境[44?46]。不同恢復年限人工草地植被分布有差異,因而土壤微生物分布特征也不相同。龍健等[25]指出,植被能夠通過凋落物影響微生物的分布,凋落物的類型對土壤養分平衡十分重要。本試驗調查樣地為禾本科植物混播草地,建植初期草地凋落物絕大部分為禾本科植物凋落物,具有較高的C/N(100/1),而隨著人工草地的恢復演替,豆科和其他類型植被逐漸出現,平衡C/N,促進草地土壤養分的進一步平衡,為微生物的生長提供更好的條件。此外,真菌能夠通過分泌胞外多糖從而提高抗逆性,但細菌卻沒有該功能[47?48],因此在草地恢復過程中真菌表現出更大優勢。但隨著草地恢復,環境得到改善,細菌也在恢復中。真菌在初期土壤穩定性形成過程中起著十分重要的作用,可通過菌絲不斷增加土壤顆粒之間的團聚性,促使土壤大團聚體穩定形成。同時在土壤有機質或凋落物分解、土壤速效養分積累中真菌表現出促進作用[49],為進一步促進其他微生物的恢復和積累創造了優良生態環境,所以隨著建植年限的增長,各類土壤微生物逐漸增多。由于人工草地凋落物中大量碳元素通過真菌作用回歸土壤,同時真菌也會加速土壤氮釋放(N2O)于大氣中[50],使土壤C/N 進一步提高,加劇了土壤肥力、營養結構的不均衡。周麗霞等[9]也在研究中指出過高的真菌比例不利于土壤綜合肥力提高,所以為避免禾本科人工草地土壤C、N 失衡,在后期管理中要適當增加氮肥維持養分平衡。
3.2 植被對人工草地土壤微生物多樣性的影響
就真核微生物而言,不同建植年限人工草地具有相同的優勢菌,但優勢菌門相對豐度具有一定差異,差異可能是不同建植年限人工草地中碳源種類決定的。如人工草地建植過程中出現的豆科植物能夠分泌脂肪酸活化土壤中難溶解的養分,為微生物提供更多氮源[5],減輕人工草地初期C/N 化合物不平衡對微生物和土壤產生的不利影響。在屬水平上,不同恢復年限草地土壤真核微生物變化特征與門水平上的變化具有相似性,隨著人工草地恢復時間變長,主要優勢屬沒有發生變化,但有害微生物屬的相對豐度卻逐漸降低,可能是因為演替過程中土壤質量的提高和一些植被根系分泌的類黃酮和酚酸類有機物,均會對有害微生物,特別是對病原微生物產生抑制作用[51?52],從而降低有害微生物的豐度。該結果與邵玉琴等[53]和楊希智等[54]的結果相似,表現為隨著草地的恢復微生物群落結構逐漸優化。但不同建植年限的人工草地土壤真核微生物結構相差很大,真核微生物恢復到穩定狀態可能還需要更長時間。
變形菌門和放線菌門微生物是不同恢復年限人工草地中的主要原核優勢菌門。這兩個門下的許多微生物在各類環境下甚至極端條件下均能生存[55?56],但優良的環境更有利于變形菌和放線菌大量積累[6],隨著人工草地演替逐漸穩定,變形菌門、放線菌門微生物豐度的增多也驗證了這一觀點。探究植被和主要真核、原核微生物的關系發現,植被特征與原核微生物優勢屬無明顯的顯著關系,但與真核微生物關系密切(表3)。植被不僅能夠通過為微生物提供不同的碳源對其群落結構和分布產生影響,也能夠通過改變土壤理化性質,間接對土壤微生物產生影響。有學者提出養分及細顆粒土壤對土壤微生物的生長有很重要的作用[57],而植被在積累養分,降低土壤粒徑上效果顯著[58?61],所以植被可通過改善土壤理化性質間接促進土壤微生物的恢復。此外土壤水分也是土壤微生物分布及多樣性的關鍵影響因子[62?64]。在干旱半干旱地區,土壤水分往往作為第一限制因子影響微生物和植被生長[7,64]。在青藏高原地區土壤水分在微生物恢復和提高微生物活性也起重要作用。研究表明,建植初期的人工草地土壤水分很低,隨著草地的演替,植被的保水能力增強,人工草地土壤含水率明顯增加[65]。本研究發現,土壤含水率低的人工草地(1,4,11 年)中微生物物種數、多樣性指數均較低。而建植18 年的人工草地中微生物物種數、多樣性指數較高。多樣性高的微生物群落表現出較高的生態功能。如演替后期變形菌門中許多具有異養光合作用菌種的出現和豐度的增加,提高碳、氮循環,增加土壤碳氮含量[1,7,41,43,50],緩解了人工草地土壤養分的失衡。此外,一些放線菌也能夠促進有機體降解,而某些變形桿菌能分泌粘性蛋白,促進土壤顆粒的粘附[57],穩定和改善土壤結構,所以微生物的恢復和活性的升高對草地土壤和植被的恢復有重要意義。
人工草地的恢復演替是植被和微生物共同作用的結果。但對比健康高寒草甸[32],人工草地中真核微生物占據主導地位,雖說原核微生物有較高的多樣性,但主要原核、真核優勢菌的相對豐度還存在較大差異,所以人工草地微生物要想恢復到健康高寒草甸水平還需要更長時間。
4 結論
隨著人工草地建植年限的增加,土壤表層微生物OTUs 數量和多樣性隨之增加,但原核微生物多樣性(除Simpson 指數)顯著高于真核微生物多樣性;不同建植年限人工草地真核、原核主要優勢門、屬一致,但相對豐度具有一定差異,其中不同年限間原核微生物相對豐度差異小,真核微生物相對豐度差異大,所以人工草地微生物群落結構還在變化,還需要更長的時間才能達到穩定狀態。人工草地的植被特征與原核微生物沒有相關性,但植被總蓋度和真核微生物OTUs 數量、優勢菌(門、屬)關系密切。總體而言,隨著人工草地建植年限增加,植被群落的恢復演替促進了土壤微生物的恢復。
