微生物共培養生產化學品的研究進展
2021-02-10 05:32:10李向來申曉林王佳袁其朋孫新曉
合成生物學 2021年6期
關鍵詞:系統
李向來,申曉林,王佳,袁其朋,孫新曉
(北京化工大學,化工資源有效利用國家重點實驗室,北京 100029)

隨著合成生物學的發展,生物合成已成為化學品制造的重要方式[1]。理論上,途徑已知的任何化學品都可通過生物合成由廉價碳源實現生產。傳統上,微生物合成化學品以單菌株培養為主,即單一細胞攜帶完整的代謝途徑。目前通過單培養已實現許多化學品的工業化生產,但在合成長途徑及復雜途徑化學品時還存在較大挑戰,生產性能往往達不到工業應用要求。代謝負擔和代謝壓力是影響生產性能的重要障礙:合成途徑與細胞生長競爭有限的資源;途徑的引入可能會打破宿主原有的物質能量代謝平衡;終產物及中間體的積累可能會產生細胞毒性。另外,由于宿主細胞缺乏內膜結構及翻譯后修飾系統,無法為某些外源酶的功能表達提供合適的細胞微環境。針對上述問題,借鑒自然界中的共生現象,研究者開發了共培養技術,通過在同一體系中培養兩種或多種細胞實現勞動分工及代謝分區。與單培養相比,共培養在減輕宿主代謝負擔[2-3]、提供合適的酶催化環境[4]以及復雜/混合/非常規底物利用[5-9]等方面具有顯著優勢,甚至可以合成通過單培養無法合成的化學品[10-11]。
1 微生物共培養減輕代謝負擔生產化學品
目標產物合成與細胞自身代謝競爭前體和能量等資源。在單一菌株中引入復雜途徑可能會導致嚴重的代謝負擔。將途徑拆分并分配至不同細胞可分擔代謝壓力。此外,各模塊可單獨進行表達優化,通過菌株比例的調節可實現模塊間的平衡。近來,在共培養減輕代謝負擔生產化學品方面取得諸多研究進展(表1)。
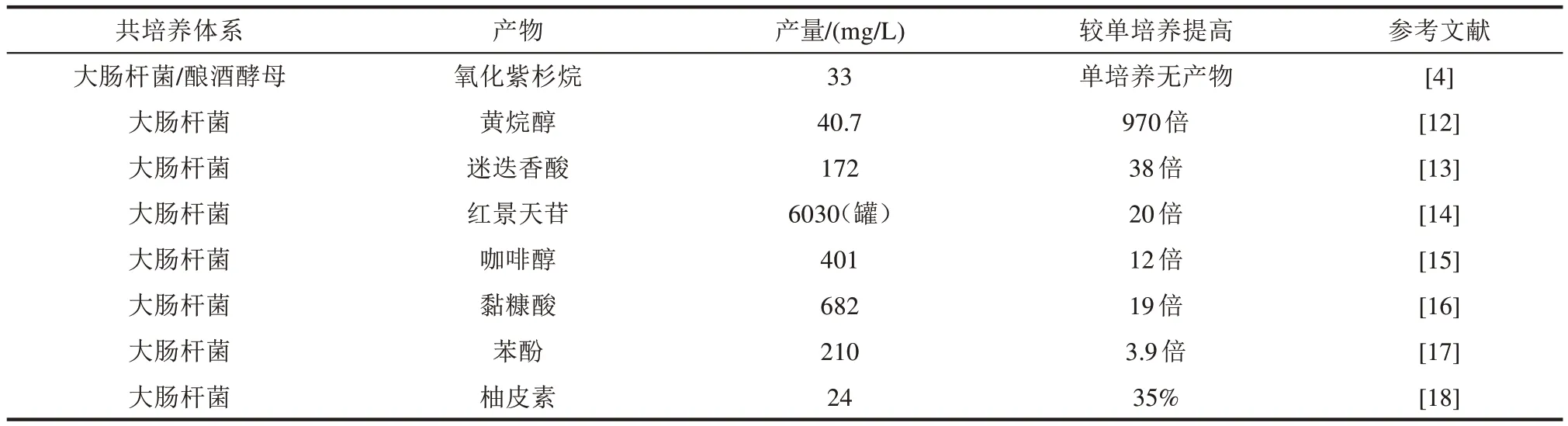
表1 共培養生產化學品的代表性研究進展匯總Tab.1 Summary of representative research progresses on microbial co-culture to produce chemicals
在生物合成黃烷醇的過程中,研究者基于對輔因子和前體的不同需求,將途徑拆分為上游丙二酰輔酶A 依賴性模塊和下游NADPH 依賴性模塊,并分配到兩個大腸桿菌菌株中。通過啟動子文庫對上游途徑進行表達優化,并對組成下游途徑的基因進行組合優化。接著,對共培養體系的誘導時間、誘導溫度、接種比例和碳源4個參數進行了優化,并基于初始優化數據建立了經驗尺度的高斯模型,預測最佳生產條件,最終產量達到40.7 mg/L,較單培養產量提高970倍[12]。
除了線性途徑,共培養策略在非線性途徑的平衡優化方面也具有突出優勢。例如,迷迭香酸(RA)是一種植物多酚類活性天然產物,其合成途徑屬于分支-匯聚型。RA 的兩種結構單元咖啡酸(CA)和丹酚酸(SAA)均來自同一前體酪氨酸,存在競爭關系;另外兩者按等摩爾比酯化生成RA,又存在協同關系。單培養策略很難達到兩個模塊之間的微妙平衡,以實現代謝資源的合理分配。為此,研究者將RA 的生物合成途徑拆分為3個模塊(CA模塊、SAA模塊以及RA模塊)并分配到3 種大腸桿菌工程菌株中。CA 模塊包含3 種酶,將4-羥基苯丙酮酸轉化為CA;SAA 模塊包含羥化和還原兩步反應,將4-羥基苯丙酮酸催化為SAA;RA 模塊先將CA 轉化為咖啡酰輔酶A,然后迷迭香酸合成酶催化咖啡酰輔酶A 和SAA 生成RA。為了增加體系的穩定性,通過改造使得CA和RA合成菌株優先利用葡萄糖生長,而SAA合成菌株利用木糖生長,優化接種比例后RA 產量達到172 mg/L,較單培養提高38倍[13]。
當單獨優化的菌株進行物理混合和共培養時,菌群比例經常發生動態變化。由于生長競爭和代謝壓力,共存狀態可能會崩潰。除了上述碳源的選擇性利用,研究者還通過建立互利共生關系來提高菌株比例的穩定性。例如,Liu 等[14]建立了交叉喂養的共培養體系用于紅景天苷的生物合成。前期的研究表明大腸桿菌單培養合成紅景天苷效率低下,大量中間體酪醇未被糖基化。為此,將其合成途徑分為酪醇生產模塊以及UDP-葡萄糖和紅景天苷生物合成模塊,并分別導入代謝木糖的苯丙氨酸缺陷菌株和代謝葡萄糖的酪氨酸缺陷菌株。通過酪氨酸和苯丙氨酸的營養共生,形成了穩定的大腸桿菌共培養系統,紅景天苷最終產量達6.03 g/L,較單培養提高20倍。
此外,共培養策略還成功應用于木質醇、黏糠酸、苯酚、白藜蘆醇等化合物的生物合成[15-23]。
2 微生物共培養利用復雜、混合及非常規底物生產化學品
聯合生物加工(CBP)旨在直接將木質纖維素生物質轉化為燃料及化學品。然而,工業生產菌株往往不具有木質纖維素降解能力,而木質纖維素降解菌株生產能力較弱或者缺乏遺傳操作工具。研究者嘗試構建可將纖維素轉化為乙醇或其他燃料的單一菌株。例如,表面展示纖維小體的釀酒酵母可利用微晶纖維素生產乙醇[24]。然而,單菌株既要分泌纖維素酶又要合成產物,代謝負擔往往較重,影響最終產量。為此,研究者嘗試共培養策略進行勞動分工,其中一種菌株分泌纖維素酶,另一菌株將水解產生的單糖轉化成產物。例如,可水解纖維素的梭狀芽孢桿菌和產乙醇的嗜熱厭氧菌混合培養,乙醇產量較單培養提高 4.4 倍[25]。Nakayama 等[26]設計嗜熱厭氧梭菌和淀粉產丁醇梭菌的共培養體系,直接由纖維素生產丁醇。然而,由于兩種菌株最適生長溫度不匹配,需要進行高溫糖化和中溫發酵兩階段培養,增加了過程能耗及復雜性。為此,Wang等[6]通過富集培養獲得了中溫條件下穩定高效降解纖維素的微生物菌群N3,并對組成菌株進行了分類鑒定,其中快生梭菌N3-2 具有最佳的纖維素降解效率,進而分別建立了N3 和N3-2 與丙酮丁醇梭菌的共培養體系,結果表明前者具有較高的丁醇生產效率,產量可達3.73 g/L。其他的應用實例還包括里氏木霉/大腸桿菌共培養產異丁醇、里氏木霉/德氏根霉共培養產富馬酸等[7,27]。
木質纖維素水解液含有葡萄糖、木糖和阿拉伯糖等單糖。葡萄糖效應的存在限制了碳源共利用。通過定向進化及表達相關轉運蛋白可實現單菌株同時利用葡萄糖和木糖[28-30]。此外,通過共培養策略也可實現碳源共利用。例如,3 種工程酵母菌株共培養可以發酵葡萄糖-木糖-阿拉伯糖的混合物,與可代謝3種糖的單菌株相比,馴化后的共培養菌株在長時間重復批次培養時具有更穩定的發酵動力學[31]。共培養策略還用于甲烷、CO 等非常規碳源的利用。例如,Hill 等[32]設計了藍細菌-甲烷氧化菌共培養體系,實現光合作用與甲烷氧化相偶聯,將溫室氣體CH4和CO2等轉化為微生物生物量,通過光照控制可以實現體系的連續穩態運行。該體系在沼氣和天然氣等混合氣體中生長良好,且與傳統的甲烷異養菌單培養相比,不需要輸入CH4/O2混合氣體,提高了過程的安全性。重要的是上述兩種細菌均可進行代謝工程改造,因此該平臺可進行擴展,以CH4為廉價原料合成各種化學品。Lee 等[33]設計了檸檬酸桿菌與卵形鼠孢菌的共培養體系用于CO 轉化。檸檬酸桿菌可通過CO 脫氫酶將 CO 和 H2O 轉化成 CO2和 H2,卵形鼠孢菌進而將CO2和H2轉化成乙酸,共培養乙酸產量較卵形鼠孢菌單培養提高82%。另外,通過自養微生物(藍細菌等)和異養微生物(細菌、酵母及真菌等)共培養可以直接通過光合作用固定CO2合成化學品[34-39]。
3 微生物共培養擴大化學品多樣性
微生物是各種活性化合物的重要來源。通過基因組測序分析后發現微生物中天然產物基因簇的數量遠遠超過實驗室條件下檢測到的化合物的數量,原因之一是單培養條件下缺乏生物或化學刺激,導致合成途徑處于沉默狀態[40]。自然條件下,微生物共生形成群落,相互之間存在物質及信息交流。研究表明共培養可顯著影響微生物代謝[41]。近年來,液質聯用等分析技術的發展促進了共培養誘導天然產物合成的研究[42]。通過共培養技術已發現包括聚酮、大環內酯及二萜等數百種新化合物[43-48]。例如,通過共培養土棲棘殼孢(Setophoma terrestris)和解淀粉芽孢桿菌(Bacillus amyloliquifaciens),Arora 等[49]分離到一種新的聚酮化合物Blennolide K。從稻黑孢霉(Nigrospora oryzae)和白囊耙齒菌(Irpex lacteus)的共培養體系中,Zhou 等[50]分離到5 種新的倍半萜烯化合物。
如上所述,共培養技術在化學品的生產和發現方面取得了重要進展。為了更好地預測及控制菌株比例,近年來研究者在調控工具開發及計算機模擬方面開展工作,以推動共培養技術的進步。
4 群體感應調控菌群比例
在微生物共培養中,成員之間的密度比例是系統功能的決定因素之一,影響到系統穩定、基因表達以及生產效率,因此控制共培養中種群密度比例的方法至關重要。由于多細胞共培養各成員之間復雜的相互作用,往往共培養環境中一個微小的變化就會引起密度比例的大幅波動。因此,想要在實際生產中構建出符合人們期望比例的或者比例能夠隨生產環境變化而變化的共培養系統還面臨巨大挑戰。目前,關于控制共培養種群密度比例的研究還比較少,最常用的手段包括調節接種比例、微流體以及固定化培養等。這些方法還難以實現群體比例的過程控制。為此,研究者嘗試利用群體感應(QS)等策略實現群體比例的自主調控(圖1)。
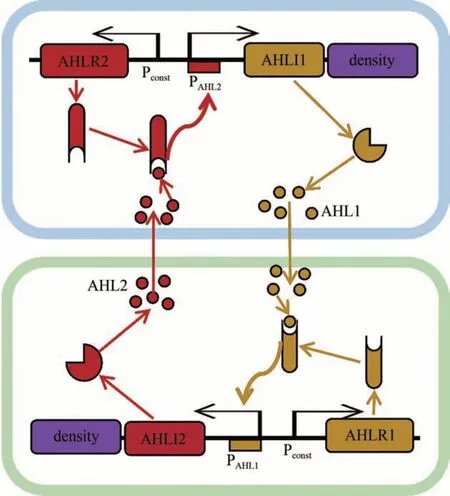
圖1 基于群體感應的共培養體系Fig.1 A quorum sensing-based co-culture system
QS 系統由于其簡單的遺傳結構和天然的多樣性,已廣泛用于工程化細胞間的通信,其中代表性的是以小分子酰基高絲氨酸內酯(AHL)為信號分子的QS 系統。AHL 由酰基高絲氨酸內酯合成酶合成,不同來源的合成酶產生不同結構的AHL信號分子,并響應不同的QS 系統。在運用QS 的微生物共培養系統中,一種微生物經過工程化設計可以合成某種特定的AHL 信號分子。該信號分子可在細胞內外環境中擴散并與對應的轉錄因子蛋白結合并形成活化的復合物,激活特定的QS 啟動子并啟動下游基因轉錄[51]。來自費氏弧菌和銅綠假單胞菌的lux 和las 系統是最常用的QS 系統,均有著靈敏的信號響應性、對同源AHL 信號分子的特異性以及與大腸桿菌的相容性,已被廣泛應用于合成生物學[52]。為了擴大QS系統的多樣化運用,tra、rpa、rhl、cin 和 esa 等新型 QS 系統得到開發[53-54]。
Chen 等[55]使用兩種不同的 QS 系統來構建“激活”菌株和“阻遏”菌株,其中“激活”菌株(rhl 系統)在rhl-AHL 信號分子存在下可激活兩種菌株中目標基因的轉錄,而“阻遏”菌株(cin 系統)在cin-AHL 信號分子存在下通過表達阻遏物LacI 同時降低兩種菌株中靶基因的轉錄。此外,cin-AHL 還將直接激活AiiA 的表達,該酶可降解上述兩種信號分子,起到負反饋作用。當兩種菌株共培養時,這兩種信號調控機制會共同在種群水平上產生耦合的正反饋和負反饋調節,并且由于AiiA 的存在,這種負反饋的引入將緊密調節阻遏物的濃度,對共培養系統的種群密度穩定具有關鍵作用。而最終的結果表明,采用這種雙QS 系統構建的共培養體系具有很強的魯棒性,與啟動子工程結合后可成功實現種群密度的動態調控。
Scott 等[56]利用 lux 和 rpa QS 系統成功設計出以兩個菌株同步裂解為動態調控核心的共培養系統。兩種菌株分別攜帶lux 和rpa 系統,并表達對應的信號分子lux-AHL 和rpa-AHL。隨著共培養菌株密度的增長,當兩個菌株中各自的AHL 分子濃度達到一定的閾值時,將分別激活對應的啟動子并促使細胞裂解蛋白X174 的表達,造成菌株裂解,以達到調控種群密度的目的。之后,未裂解的菌株將會繼續生長,并在達到下一次閾值時開始裂解循環。當維持兩種信號分子處于低濃度時,共培養系統將發生恒定裂解,即生長與裂解持平,而種群密度將保持在特定的比例。
McCardell等[57]設計大腸桿菌共培養體系,通過兩個正交的QS 系統控制毒素的產生,以維持特定的種群比例。通過誘導劑IPTG 或aTc 的誘導,兩種菌株產生不同的AHL 分子,在激活自身CcdB的表達(一種細菌毒蛋白,可促使DNA 裂解并阻止轉錄)的同時還將激活另一菌株中CcdA 的表達(一種抗毒素,可抑制CcdB)。在人工添加相應誘導劑的情況下,高密度菌株表達CcdB 抑制自身生長,同時促進另一菌株表達CcdA 而繼續正常生長。通過改變IPTG 和aTc 誘導劑的濃度,可以調節兩種菌株的密度達到特定比值。
AI-2 是一種存在于許多細菌中的自動誘導分子,能廣泛響應細菌的種群密度[58]。Stephens等[59]基于 AI-2 的誘導特性,在E.coli中構建了一種新型的共培養系統,并成功實現種群密度比例的動態調控以及預測。該系統首先構建了一個能感應AI-2 并能將其轉化為正交QS 信號AI-1 的菌株。產生的AI-1信號自由擴散進第2種菌株,并激活PtsH 蛋白表達,通過增強葡萄糖轉運促進細胞生長,通過動態控制第2種菌株生長速率達到調節菌群比例的目的。最后,以初始AI-2 的不同響應濃度為函數的主要變量,創建出能預測最終種群比例的數學模型,并在該模型指導下,證明這種新型的調控系統在一定種群比例范圍內的可靠性。
5 計算機模擬工具預測菌群動態變化
生物系統具有復雜性,通常需要計算模型指導代謝網絡的設計改造,以使工程菌株按照設計執行功能。共培養體系的功能分析可以借助典型的微生物組工具,包括細胞群落特定組分分析、宏基因組測序、宏轉錄組學和宏蛋白質組學等。此外,微流體細胞分選技術可以分離單個物種[60],用于研究它們的DNA、RNA 或蛋白質特征。同時,對代謝物的實時監測還可以研究共培養組分的整體動態變化。結合這些數據分析手段,越來越多的微生物群落的計算機模擬工具被成功開發。
基因組規模模型(GSMs)是一種基于全基因組序列的模擬方法。該方法將目前所有已知的調控代謝的反應合并到合適的數學分析網絡中,再結合菌株生產目標以及相關的約束條件進行共培養模擬,用以預測菌株在不同底物上的生長情況、特定基因敲除對細胞代謝的影響以及菌株間可能的相互作用等微生物功能。目前,GSMs 已被應用于微生物共培養模式及其相互作用機制的研究。例如,Koch等[61]利用化學計量模型預測微生物群落的組成,并應用于沼氣生產過程。通過模擬得到3種微生物分別在單培養、雙培養和三培養情況下的全部化學反應,并利用分級優化的方法對生長速率、底物利用率以及產量進行逐級優化,成功預測了在不同底物下的種群組成、不同生長速率對最大甲烷產量的影響以及實現最大甲烷產量的最佳營養成分。更重要的是,GSMs 操作簡便,易將單培養模型推及雙培養甚至三培養,有利于實現能量、物質的最佳化共用以及多物種間復雜相互作用的研究。結合基因表達或翻譯數據作為反應限制條件, 這些模型還可用以捕獲DNA→RNA→蛋白質的信息傳遞,已被用于研究未經人工培養的共生菌并鑒定其代謝缺陷[62-63]。
通量平衡分析(FBA)是一種在基因組規模模型基礎上建立的模擬代謝網絡的方法。該方法只需知道共培養中每個反應的化學計量系數,就可以對微生物群落中幾乎所有的代謝相互作用進行建模,并使其適應特定的環境變化。此外,FBA 還可用于預測在不同碳源中菌株的生長速度和某種代謝產物的生產速度。例如,Stolyar 等[64]研究了以乳酸為唯一碳源和能源的脫硫弧菌和馬氏甲烷球菌的共生系統,通過FBA 模型將不同代謝物間的相互關系和代謝物在化學反應中的通量以一系列的線性方程表示,以參與甲烷生產的170 個反應為模型的核心反應,并考慮不同共培養環境和物種遺傳干擾帶來的影響,將建模結果與在連續生物反應器中共培養數據進行比較,發現該模型可準確預測群落組成、代謝通量以及生長表型。
然而,FBA 是一種偽穩態模型系統,其模型較為理想化。在處理多微生物群落的復雜時空動態變化時,還需要采用動態通量平衡分析(dFBA)。dFBA 是在FBA 的基礎上充分考慮共培養體系中各種可能的變化后建立的。其可以模擬各個穩態模型在不同共培養時間階段的相互作用。此外,在dFBA 中,通過監測關鍵中心代謝產物可以驗證模型準確性。目前已經開發了針對多種微生物共培養的dFBA。例如,在纖維素分解梭菌和產溶劑丙酮丁醇梭菌的共培養模型中,dFBA 被用來預測動態菌群比例、群落間相互作用以及基因表達水平[65]。常微分方程模型被用來模擬不同的共培養相互作用類型(互利共生、偏利共生等)。例如,Kong等[66]以營養成分、兩種菌株及各自所產生的相互作用分子為主要變量,結合生長速率、相互作用強度等因素,再通過模塊化途徑重構與計算機模擬,構建了6種不同作用方式的乳酸乳球菌雙菌株共培養體系,進一步由簡至繁,推測出三菌株以及四菌株的共培養模型,并成功預測其種群動態、種群行為以及相關的動力學特征。另外,研究者基于Monod 微分方程開發了計算機模型,成功預測以碳源(有機酸)和氮源(銨)營養共生的大腸桿菌和沼澤紅假單胞菌的最佳發酵條件,并形成了穩定的共培養體系[67]。模型結果表明,大腸桿菌產生的有機酸(如甲酸、乙酸和乳酸等)具有抑制作用,將會降低碳的利用效率和破壞種群平衡。
6 挑戰和展望
隨著合成生物學的發展,微生物共培養已成為化學品生物合成的新方法。利用該方法,不僅可以減輕代謝負擔實現復雜化合物的合成,還可以充分發揮不同物種的優勢和能力,利用低劣生物質以提高目標產品經濟性。雖然微生物共培養已經展現出許多優勢,但目前仍存在一些問題和挑戰。首先,共培養體系是一個動態變化的系統,難以實現長期的穩定。由于成員生長速度不同或者異源物種不兼容,通常出現某一成員完全取代另一成員的情況,導致生產性能降低甚至共培養系統完全破壞。雖然已有許多研究通過QS 系統來保持種群的穩定性,但是這些系統在進行長時間的發酵時也表現欠佳。目前,保持種群穩定性的最有效方法是建立個體成員間牢固的互利互惠或共生關系。其次,共培養體系各成員間可控的種群比例也是一個值得關注的問題。可控的種群比例對于生產途徑中的代謝通量平衡至關重要,可實現目標產物的最優化生產。然而,共培養成員間的相互作用是復雜多樣的,往往一個微小的變化都會引起種群比例的巨大變化,從而導致通量平衡被破壞。盡管以前的研究通過表達裂解蛋白[56]、毒素蛋白[57]或其他負反饋抑制機制[55]來阻止細菌生長從而調控種群比例,但是這些研究不僅將產生額外的代謝壓力,這種抑制性的種群比例調控方式還將導致巨大的生物質浪費。同時,隨著共培養體系中所包含的成員越多,解決這個問題的難度將越來越大。目前,解決這個問題最可能有效的方法是通過QS 系統或其他常規小分子響應器來正向調控共培養體系成員的生長,例如Stephens 等[59]的研究,而非阻止或殺死。此外,中間體的轉運也是一個關鍵的問題[68]。途徑的拆分意味著中間體需要從一個菌株轉運到另一個菌株。但是某些化合物不能自由通過細胞膜或者不能被第二菌株攝取。面對這種情況,通常需要表達額外的轉運蛋白。最后,異源菌株的共培養是面臨的另一挑戰。不同種屬的菌株有著不同的生物合成能力,例如,釀酒酵母具有完善的蛋白表達修飾機制和豐富的細胞內膜,是表達細胞色素P450s 的理想宿主[69],而枯草芽孢桿菌可利用自身合成的生物表面活性劑來提高疏水有機化合物的生物利用度,其已被廣泛應用于原油的降解等[70-71]。因此,利用異種菌株的共培養來生產化學品前景廣闊,但不同種菌株的生長生產條件差異顯著,將造成物種間不兼容。未來,微生物共培養的發展將受益于合成生物學相關技術的進展,包括基因組工程、宏蛋白質組學、宏代謝組學以及CRISPR/Cas 工具等。微生物共培養未來發展的一個方向是更廣泛地兼容多物種共培養,以利用不同物種的生物合成能力。這對于植物和真菌的天然產物生物合成具有特別意義,因為在共培養中使用真核生物對于植物和真菌天然產物途徑酶的功能性表達非常有益。未來發展的另一個方向是開發針對萜類化合物和生物堿等結構更復雜的化合物的高效合成共培養策略。總體而言,通過一系列元件、工具及模型的開發,上述問題和挑戰將逐步得到解決,未來將可設計復雜穩定可控的共培養體系應用于化學品的高效生產。
致謝:感謝國家重點研發計劃項目(2018YFA0901800)及北京化工大學雙一流項目(XK1802-8,XK1902)對本研究的支持。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
