青藏高原黑果枸杞花青素對人肝癌細胞增殖和自噬的影響
2021-02-01 10:19:02王夢杰李玉林劉嘉華張龍飛阿孝蓉周佳俊柴沙駝
天然產物研究與開發 2021年1期
關鍵詞:肝癌
王夢杰,李玉林,劉嘉華,張龍飛,阿孝蓉,周佳俊,吳 華*,柴沙駝
1青海大學農牧學院;2中國科學院西北高原生物研究所;3青海大學畜牧獸醫科學院,西寧 810000
肝臟是人體內最主要的代謝器官之一,具有合成分泌型蛋白質、去氧化和儲存肝糖等功能。肝癌是全球第六大常見癌癥。與其他實體癌相比,過去十多年來,世界范圍內肝癌的發病率和死亡率都呈現增長趨勢[1]。在各類癌癥的發病率和死亡率中,肝癌的發病率和死亡率均位于前十位(發展中國家中發病率85%,居不同癌種第一),中國作為最大的發展中國家,肝癌發病總數占全球肝癌病人總數的50%[2],肝癌已經成為國民健康的主要威脅者之一。在多種導致肝癌的因素中,人類飲食習慣和生活習慣的失調是重要原因之一。在人們崇尚綠色天然生活觀念的影響下,利用天然食品中的抑癌因子來有效預防和干預癌癥的發生發展已獲得人們的重點關注。
黑果枸杞(LyciumruthenicumMurray)是茄科枸杞屬植物,漿果呈現為球形,果實成熟后顏色呈紫黑色,果實無毒且具有甜味,是一種尚待開發的野生植物資源[3]。黑果枸杞主要分布于我國西北地區,因其富含花青素而被譽為“沙漠黑珍珠”和“花青素之王”[4]。花青素(anthocyanidin,又稱花色素),屬類黃酮化合物,具有抗腫瘤、抗炎、降血糖血脂、護肝以及很強的抗氧化能力等多種生理功能[4-6]。在食品開發和醫學研究方面具有較高的研究價值。基于花青素的生理活性和保健作用,黑果枸杞儼然已經成為青海省的特色經濟植物,并在醫療保健領域獲得了廣泛關注。然而,目前對黑果枸杞花青素的研究還停留在黃酮、多糖等成分的抗氧化活性上,尚未見黑果枸杞花青素對人肝癌HepG2細胞自噬作用影響機制的相關報道。基于此,本試驗主要針對黑果枸杞花青素對人肝癌HepG2細胞免疫活性的影響展開相關研究,為青藏高原黑果枸杞的深度開發利用提供一定的理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 試驗材料
細胞系:人肝癌細HepG2細胞由中國科學院細胞庫提供。
黑果枸杞花青素:購自青海金麥杞生物科技有限公司(花青素:64.4%;灰分:0.6%;蛋白質:6%;糖類:1.32%;氨基酸:3.56%)。
1.1.2 主要試劑配制
PBS:將PBS干粉倒入容量瓶,向容量瓶中加入1 L超純水,并用超純水反復沖洗粘在袋壁上的粉末并移至容量瓶,充分攪拌至PBS干粉完全溶解后將其定容至2 L,調節PBS溶液的酸堿度為7.2~7.4。而后將PBS溶液轉移至玻璃容器中進行高壓蒸汽滅菌,待溶液冷卻至室溫后,用封口膜密封并放至4 ℃冰箱低溫保存待用。
DMEM培養基:將DMEM粉末溶于超純水和FBS,用超純水定容至1 L,控制FBS的終濃度為10%。加入適量的碳酸氫鈉,調節pH值為7.2~7.4,濾菌后用封口膜密封并放至4 ℃冰箱低溫保存。
花青素溶液:稱取2 mg花青素溶于10 mL DMEM培養基中,配制得到花青素濃度為200 μg/mL的溶液,濾菌后用封口膜密封并避光低溫條件下保存。
3-MA(3-methyladenine)自噬抑制劑配制:精確稱量20 mg 3-MA,加入3.36 mL PBS緩沖液配制成濃度為20 mM的溶液,濾菌后用封口膜密封置于-20 ℃冰箱保存。
1.1.3 試劑與設備
DEPC(西安赫特);DMEM、胰酶、雙抗(Gibco公司);DMSO(山東德彥化工有限公司);EDTA(Solarbio公司);FBS、PBS、Promega反轉錄試劑盒、SYBR Green I熒光染料、SDS、無水乙醇、氯仿、異丙醇(西寧寶信生物科技有限公司);EdU試劑盒(廣州瑞博生物科技有限公司);超純水、Aglient1260高效液相色譜儀(Aglient公司);X500R ESI-Q-TOF質譜儀(SCIEX公司);乙腈、色譜純、凈化工作臺(江蘇通凈凈化設備有限公司);CO2恒溫培養箱(美國NUAIRE公司);水平離心機(英國Dynamica公司);酶標儀(無錫華衛德朗儀器有限公司);熒光顯微鏡(日本O-lympus公司);熒光定量PCR儀器、蛋白印記檢測系統(美國Bio-Rad公司);Western blot轉膜儀、電泳儀、電源(購自北京六一生物科技公司);搖床(美國Scilogex公司)。
1.2 試驗方法
1.2.1 黑果枸杞花青素提取物液質分析條件
色譜條件:Phenomenex Luna-C18柱(250 mm×4.6 mm,5 μm);流動相A(1 000 mL超純水,0.1%甲酸),流動相B(乙腈)。梯度梯度洗脫程序(0~10 min,流動相B:15%;10~30 min,流動相B:15%→40%,流速1.0 mL/min;柱溫25 ℃,檢測波長254 nm)。
質譜條件:采用TurbolonSpray離子源,氣簾氣30 psi,離子源載氣150 psi,離子源載氣260 psi,離子源溫度550 ℃;IDA正離子模式,噴霧電壓5 500 V;CAD載氣7 psi,一級TOF-MS質量掃描范圍50~1 500 Da,誘導碰撞電壓10 V;二級TOF-MS質量掃描范圍50~1 500 Da,誘導碰撞電壓45 V。
1.2.2 細胞培養
1.2.2.1 細胞的常規培養
人肝癌HepG2細胞培養24 h,棄去舊的培養基,加入5 mL新的DMEM培養液十字混勻后放入5% CO2,37 ℃的培養箱中繼續培養。
1.2.2.2 細胞計數
向血球計數板中滴加10 μL預先混好的人肝癌HepG2細胞懸液,在培養箱內靜置培養3 min后在顯微鏡下觀察計數,根據觀察得到的細胞數統計計數板中四大格細胞的總數為A,計算細胞數:實際細胞數=A/4×10×1 000×稀釋倍數。
1.2.3 CCK-8法檢測黑枸杞花青素對人肝癌HepG2細胞活率的影響
將人肝癌HepG2細胞以1×106個/mL的密度接種于96孔板中(每孔100 μL),細胞培養貼壁24 h后用黑果枸杞花青素誘導細胞,控制黑果枸杞花青素的終濃分別為0、12.5、25、50、100 μg/mL,并分別作用于人肝癌HepG2細胞12、24、36、48 h。在各時間點前的3 h,每孔加10 μL CCK-8溶液,繼續孵育3 h,用酶標儀測定其450 nm處的OD值,每組設3個重復。計算公式:細胞活力=(OD實驗組-OD對照組)/(OD對照組-OD空白組)×100%。
1.2.4 EdU熒光標記法檢測黑枸杞花青素對人肝癌HepG2細胞增殖的影響
將人肝癌HepG2細胞以1×106個/mL的密度接種于96孔板中(每孔100 μL),細胞培養24 h。設花青素刺激組濃度分別為0 μg/mL和25 μg/mL。0 μg/mL組中加入DMEM培養基正常培養人肝癌HepG2細胞;黑果枸杞花青素刺激組中加DMEM培養基并用控制終濃度為25 μg/mL的花青素誘導人肝癌HepG2細胞。每組設3個復孔,每孔終體積為300 μL,培養24 h。
首先制備出適量的濃度為50 μM的EdU培養基;每孔中均加入100 μL EdU培養基,在培養箱內靜置培養2 h后棄去培養基。用PBS清洗未貼壁的細胞;每孔加50 μL細胞固定液,恒溫培養30 min后棄去固定液;加50 μL 2 mg/mL甘氨酸,脫色搖床溫育5 min后棄去甘氨酸溶液;加入100 μL PBS,脫色搖床溫育5 min后PBS清洗5 min;最后加入100 μL的滲透劑,溫育10 min后用PBS清洗5 min。加100 μL Apollo染液,避光室溫孵育30 min后棄去染液;加100 μL滲透劑,脫色搖床清洗2次,每次10 min,而后倒掉滲透劑;加100 μL甲醇,清洗2次,每次5 min,再用PBS清洗5 min。加100 μL的DAPI反應液,在避光、室溫條件下孵育30 min后倒掉染色液;加100 μL PBS清洗細胞;觀察人肝癌HepG2細胞的染色情況。
1.2.5 細胞劃痕試驗測定測黑果枸杞花青素對人肝癌HepG2細胞遷移的影響
每隔0.5 cm在6孔板背后劃一道橫線,每孔接種1×106個細胞并且至少穿過5條線。培養24 h后用槍頭沿著橫線在細胞層上進行劃痕。用PBS清洗細胞3次,洗去未貼壁和劃線是脫落的細胞。控制黑果枸杞花青素刺激組加入終濃度分別為0、25 μg/mL,培養24 h后在顯微鏡下觀察細胞。
1.2.6 Western blot檢測增殖和自噬相關基因的蛋白表達
細胞分組設為對照組(0 μg/mL)、花青素組(25 μg/mL)、3-MA(5mM)組和花青素(25 μg/mL)+ 3-MA(5 mM)組。細胞5% CO2,37 ℃培養24 h后收集細胞。
用蛋白裂解液提取細胞總蛋白;BCA法測定蛋白濃度,調整蛋白濃度,加入適量2×SDS上樣緩沖液,煮沸5 min,-20 ℃保存備用。配制12% SDS-PAGE分離膠和5% SDS-PAGE濃縮膠,100 V,電泳2 h。根據蛋白Marker,4 ℃,200 mA轉移2~2.5 h,將目的條帶轉移到PVDF膜上。用5%脫脂牛奶室溫封閉2 h或4 ℃過夜。TBST 溶液漂洗膜3次,8 min/次,一抗孵育2 h。TBST溶液漂洗膜3次,8 min/次,二抗孵育1 h。最后用Bio-Rad Chemi Doc XRS+曝光系統檢測目的蛋白。
1.2.7 RT-PCR檢測Hippo通路和自噬相關基因表達
細胞分組設為對照組(0 μg/mL)和花青素(25 μg/mL)組,細胞培養24 h后分別用TRIZOL法和紫外線分光光度計提取總RNA和測定RNA濃度。每組各取1 500 ng總RNA,按照Promega試劑盒的說明書mRNA進行反轉錄為cDNA。反轉錄反應體系為20 μL,5×Prime Script Buffer 2(for Real Time)4 μL,Total RNA 2 μL,RNase Free ddH2O 14 μL,將體系置于PCR儀中42 ℃ 15 min;95 ℃ 3 min后獲得cDNA轉至-20 ℃長期保存。采用SYBRGreenl法以cDNA為模板對PCR進行實時熒光定量檢測,反應體系為20 μL,SYBR Premix Ex Taqll 10 μL,PCR Forward Primer 0.8 μL,PCR Reverse Primer 0.8 μL,cDNA 2 μL,ddH2O 6.4 μL,95 ℃預變性15 min,95 ℃變性10 s,60 ℃退火20 s,72 ℃延伸20 s(40次循環)。試驗每組均重復3次,根據公式2-ΔΔCt計算所測基因的相對表達量。
使用上海生物工程有限公司設計合成引物如下:
1.3 數據分析
每組試驗均重復3次,用SPSS19.0采用單因素方差分析和多重比較對試驗數據進行處理。
2 結果與分析
2.1 黑果枸杞花青素成分分析
經HPLC-Q-TOF-MS分析,從黑果枸杞花青素中確認出五種花青素化合物,總含量為64.4%。其中,C49H50O28,矮牽牛素-3-O-蕓香糖(反式對香豆酸-7-O-葡萄糖苷)-5-O-葡萄糖苷,其分子量為1 095.983,出峰時間為5.225,含量為4.2%;C43H49O23,矮牽牛素-3-O-蕓香糖(反式對香豆酸)-5-O-葡萄糖苷,其分子量為933.842,出峰時間為16.952,含量為2.6%;C44H51O24,矮牽牛素-3-O-蕓香糖(阿魏酰)-5-O-葡萄糖苷,其分子量為963.868,出峰時間為17.872,含量為47.6%;C43H49O24,矮牽牛素-3-O-蕓香糖(咖啡酰)-5-O-葡萄糖苷,其分子量為949.841,出峰時間為18.390,含量為6.4%;C44H51O23,錦葵素-3-O-蕓香糖(反式對香豆酸)-5-O-葡萄糖苷,其分子量為847.864,出峰時間為18.945,含量為3.6%(見表2)。
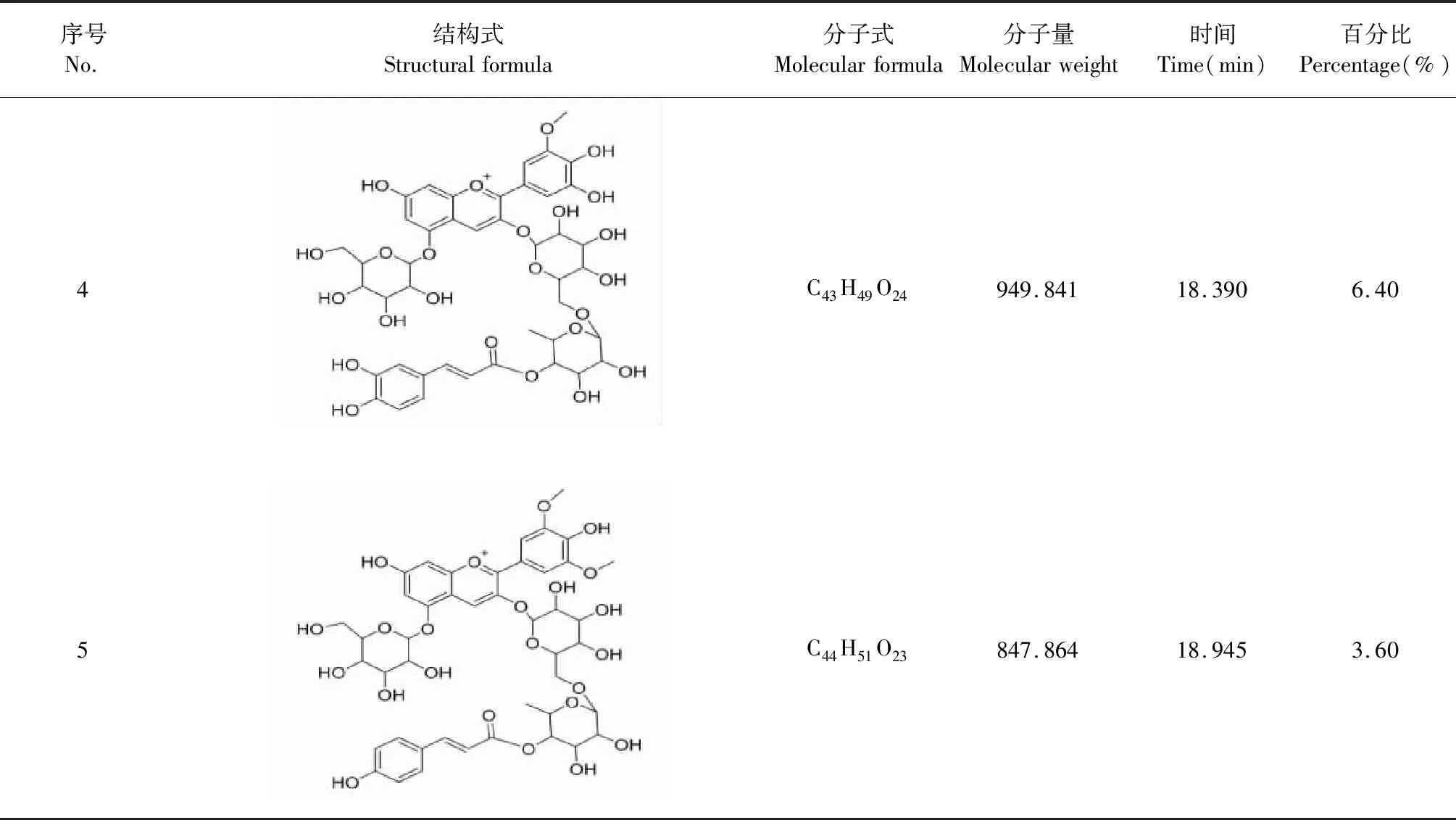
續表2(Continued Tab.2)
2.2 黑果枸杞花青素對人肝癌HepG2細胞的增殖和遷移作用的影響
2.2.1 黑果枸杞花青素對人肝癌HepG2細胞增殖作用的影響
與0 μg/mL濃度組相比,濃度為12.5、25和50 μg/mL的黑枸杞花青素作用于人肝癌HepG2細胞12 h后,人肝癌HepG2細胞的活力均顯著降低(P<0.05),濃度為25 μg/mL的黑枸杞花青素在作用于人肝癌HepG2細胞24 h后,人肝癌HepG2細胞的活力極顯著降低(P<0.01);此外,濃度為50 μg/mL的黑枸杞花青素刺激人肝癌HepG2細胞24 h后,人肝癌HepG2細胞的活力也呈顯著降低趨勢(P<0.01)。

圖1 黑果枸杞花青素對人肝癌HepG2細胞活率的影響Fig.1 The effect of L.ruthenicum anthocyanidin on HepG2 cells viability注:相同字母表示差異不顯著P>0.05,不同字母表示差異顯著,其中小寫字母表示P<0.05,大寫字母表示P<0.01。Note:The same letter indicates no significant difference P > 0.05,but different letters mean significant difference,in which the lower case letter means P < 0.05,and the capital letters means P < 0.01.
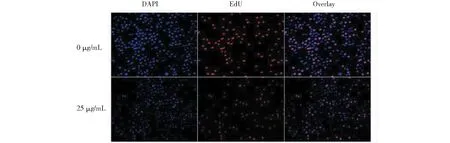
圖2 黑果枸杞花青素對人肝癌HepG2細胞增殖的影響Fig.2 The effect of L.ruthenicum anthocyanidin on HepG2 cells proliferation
與25 μg/mL濃度組相比,濃度為12.5 μg/mL的黑枸杞花青素作用24 h時對人肝癌HepG2細胞活力的抑制作用極顯著降低(P<0.01);濃度為50 μg/mL的黑枸杞花青素作用24 h和36 h時對人肝癌HepG2細胞活力的抑制作用均顯著降低(P<0.05);此外,濃度為100 μg/mL的黑枸杞花青素作用24 h和36 h時對人肝癌HepG2細胞活力的抑制作用極顯著降低(P<0.01),濃度為100 μg/mL的黑枸杞花青素作用12 h時對人肝癌HepG2細胞活力的抑制作用也顯著降低(P<0.05)(圖1)。其中,以濃度為25 μg/mL的黑枸杞花青素作用24 h時對人肝癌HepG2細胞活力的抑制作用最為顯著,因此篩選出黑枸杞花青素的最佳作用濃度和最佳作用時間分別為25 μg/mL,24 h。25 μg/mL黑枸杞花青素刺激人肝癌HepG2細胞24 h后,EdU熒光染色結果發現,與對照組相比,活細胞數減少(圖2中藍色熒光),新增殖的細胞數也明顯減少(圖2中紅色熒光),人肝癌HepG2細胞的總數也減少(圖2結合藍色和紅色熒光)。
2.2.2 黑果枸杞花青素對人肝癌HepG2細胞遷移作用的影響
細胞劃痕的結果表示,黑果枸杞花青素處理24 h后,0 μg/mL濃度組細胞連接緊密,成團生長,又重新鋪滿劃痕區;25 μg/mL濃度組的細胞在劃痕區新增細胞相對較少,細胞的增殖、貼壁和遷移能力被顯著抑制(見圖3)。
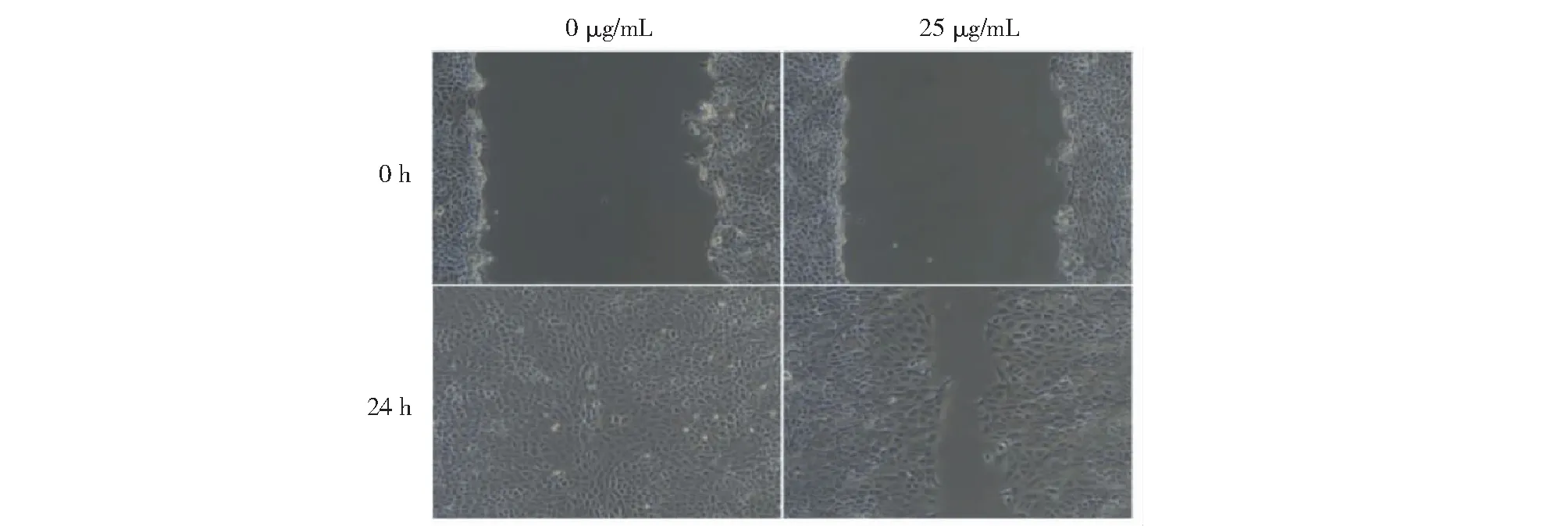
圖3 黑果枸杞花青素對人肝癌HepG2細胞遷移的影響Fig.3 The effect of L.ruthenicum anthocyanidin on migration in HepG2 cells
2.2.3 黑果枸杞花青素對人肝癌HepG2細胞關鍵增殖因子在mRNA和蛋白水平表達的影響
與對照組相比,增殖關鍵因子LATS1和MOB1在黑果枸杞花青素組細胞中的mRNA水平的表達極顯著升高(P<0.01),LATS2在mRNA水平的表達顯著升高(P<0.05),YAP在mRNA水平的表達顯著降低(P<0.05)(圖4)。花青素組CDK4的蛋白表達極顯著低于對照組(圖5,P<0.01)。
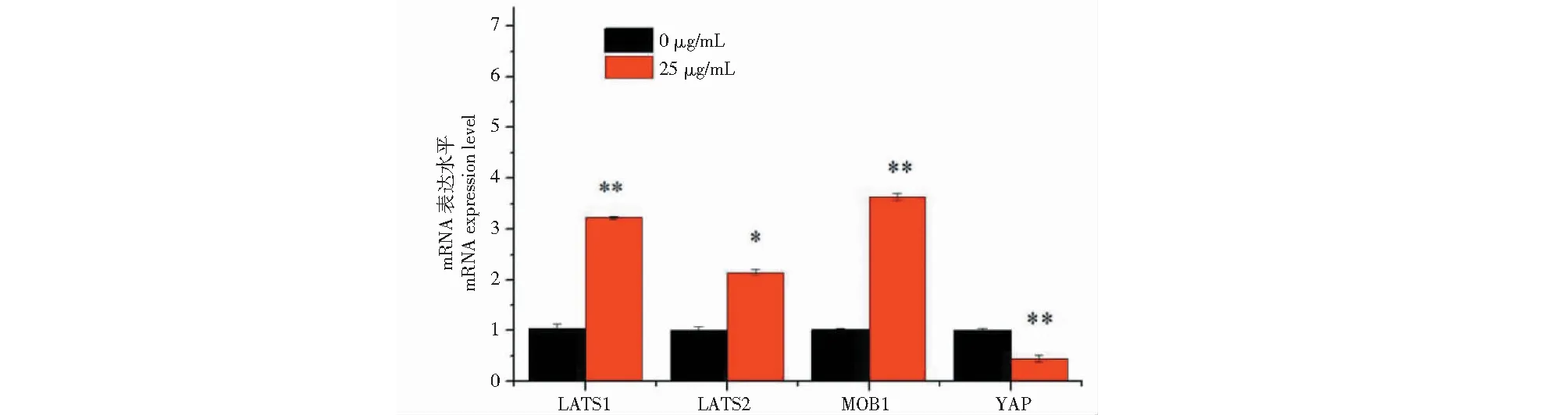
圖4 黑果枸杞花青素調控人肝癌HepG2細胞增殖因子LATS1、LATS2、MOB1和YAP在mRNA水平的表達Fig.1 L.ruthenicum anthocyanidin regulates the expression of the proliferation factors LATS1,LATS2,MOB1 and YAP at mRNA level in HepG2 cells注:與對照組比較,*P<0.05,**P<0.01。Note:Compared with control,*P<0.05,**P<0.01.
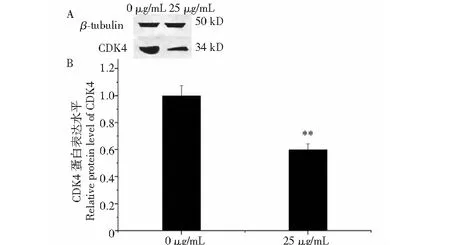
圖5 黑果枸杞花青素調控人肝癌HepG2細胞CDK4在蛋白水平的表達Fig.5 L.ruthenicum anthocyanidin regulates the expression of CDK4 at protein level in HepG2 cells注:與對照組比較,**P<0.01。Note:Compared with control, **P<0.01.
2.3 黑果枸杞花青素對人肝癌HepG2細胞自噬作用的影響
與對照組相比,黑果枸杞花青素組極顯著升高Beclin-1、LC3-Ⅱ和AMPK的mRNA水平表達(P<0.01,圖6)。與對照組相比,25 μg/mL花青素可極顯著顯著下調p-mTOR(P<0.01),并極顯著上調LC3-Ⅱ和p-AMPK的蛋白表達(P<0.01);3-MA組極顯著上調 p-mTOR (P<0.01),并極顯著下調p-AMPK和LC3-Ⅱ的蛋白表達(P<0.01);花青素+3-MA組可極顯著上調p-mTOR(P<0.01),并極顯著上調p-AMPK和LC3-Ⅱ的蛋白表達(P<0.01)。與花青素組相比,3-MA組可極顯著上調p-mTOR(P<0.01),并極顯著下調LC3-Ⅱ和p-AMPK的蛋白表達(P<0.01);花青素+3-MA組可極顯著下調p-mTOR(P<0.01),并顯著上調LC3-Ⅱ 和p-AMPK(P<0.05)的蛋白表達(見圖7)。
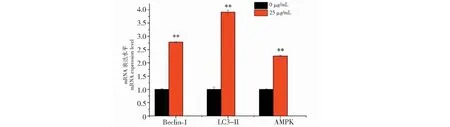
圖6 黑果枸杞花青素對人肝癌HepG2細胞自噬因子Beclin-1、LC3-II和AMPK在mRNA水平表達的影響Fig.6 The effect of L.ruthenicum anthocyanidin on autophagy factors Beclin-1,LC3-II and AMPK expression at mRNA level in HepG2 cells注:與對照組比較,**P<0.01。Note:Compared with control,**P<0.01.
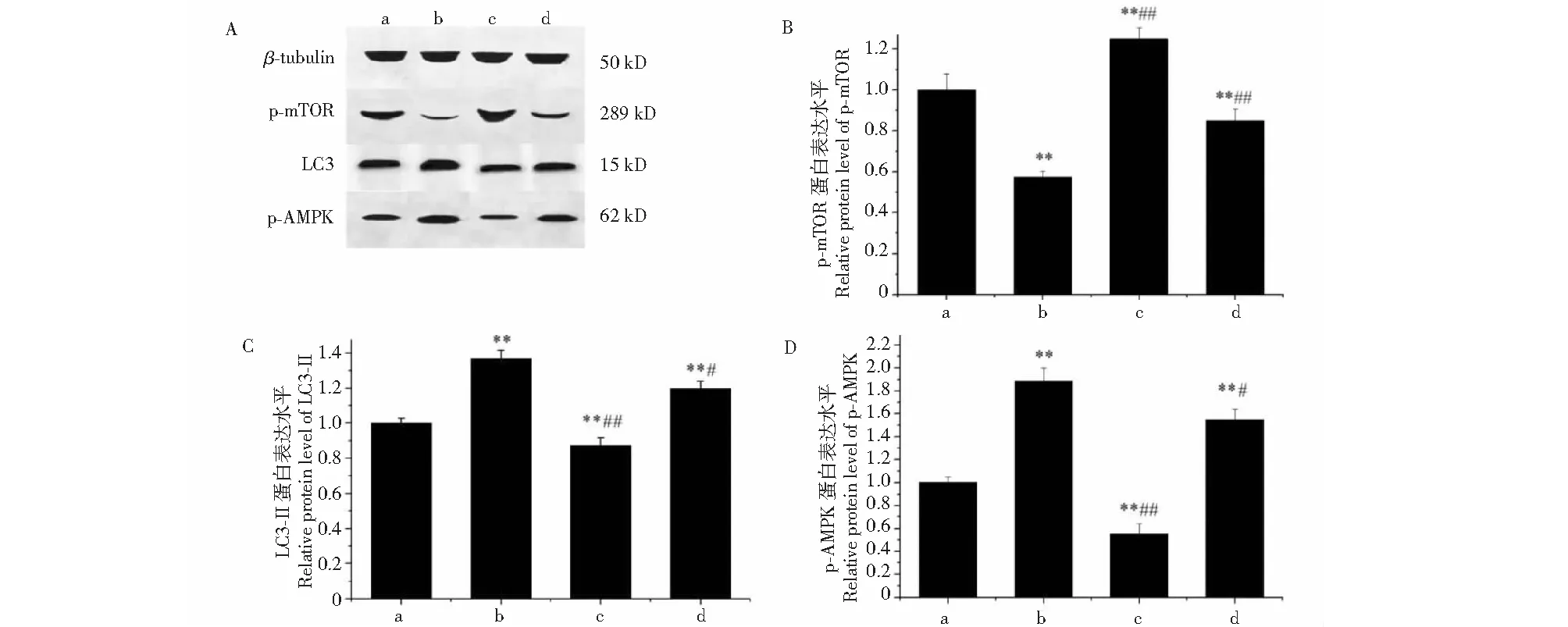
圖7 黑果枸杞花青素和3-MA對人肝癌HepG2細胞自噬相關因子p-mTOR、LC3-II和p-AMPK在蛋白水平表達的影響Fig.7 The effect of L.ruthenicum anthocyanidin and 3-MA on protein expression level of autophagy factors p-mTOR,LC3-II and p-AMPK in HepG2 cells注:a為對照組(0 μg/mL);b為花青素組(25 μg/mL);c為3-MA組(5 mM);d為花青素(25 μg/mL)+3-MA組(5 mM)。A.p-mTOR、LC3-II和p-AMPK Western blot電泳圖;B.p-mTOR蛋白相對表達量;C.LC3-II 蛋白相對表達量;D.p-AMPK蛋白相對表達量。與對照組(0 μg/mL)相比,*P<0.05,**P<0.01;與花青素組(25 μg/mL)相比,# P<0.05,##P<0.01。Note:a indicates the control group (0 μg/mL);b indicates the group induced with L.ruthenicum anthocyanidin (25 μg/mL);c indicates the group induced with 3-MA (5 mM);d indicates the group induced with L.ruthenicum anthocyanidin (25 μg/mL)+3-MA (5 mM).A.p-mTOR,LC3-II and p-AMPK Western blot figure;B.Relative expression of p-mTOR;C.Relative expression of LC3-II;D.Relative expression of p-AMPK.Compared with control,*P<0.05,**P<0.01;Compared with L.ruthenicum anthocyanidin group (25 μg/mL),#P<0.05,##P<0.01.
3 討論
3.1 黑果枸杞花青素成分分析
常見的花青素主要有6種,分別為天竺葵色素、矢車菊色素、飛燕草色素、芍藥色素、矮牽牛色素和錦葵色素[7]。而花青素在自然界中主要以糖苷化和酰基化的形式存在,以不同的結合方式形成了不同種類的花色苷,因此,通過分析花色苷來確定花青素成分是一種有效的分析方法[8]。目前,采用花青素含量的測定較為普遍的方法為高效液相色譜法以及紫外分光光度計法[7,8]。本試驗利用HPLC-ESIQ-TOF法,流動相采用水相(0.5%甲酸)和乙腈相梯度洗脫可獲得較好的分離效果和質譜響應峰值。根據高分辨質譜對分子量和同位素峰型匹配的精確性,可以確定物質的分子式,再結合二級質譜碎片信息和黑果枸杞化學成分的相關文獻報道,從黑果枸杞花青素的提取物中確認出5種衍生自矮牽牛色素和錦葵色素的花色苷,分別為矮牽牛素-3-O-蕓香糖(反式對香豆酸-7-O-葡萄糖苷)-5-O-葡萄糖苷、矮牽牛素-3-O-蕓香糖(反式對香豆酸)-5-O-葡萄糖苷、矮牽牛素-3-O-蕓香糖(阿魏酰)-5-O-葡萄糖苷、矮牽牛素-3-O-蕓香糖(咖啡酰)-5-O-葡萄糖苷和錦葵素-3-O-蕓香糖(反式對香豆酸)-5-O-葡萄糖苷,且其總含量為64.4%,說明HPLC-ESIQ-TOF分析實現了復雜中藥成分快速分離分析,為黑果枸杞微量成分分析提供了一種有效分析方法,其他化合物的化學結構則還需進一步研究。
3.2 黑果枸杞花青素對人肝癌HepG2細胞增殖作用的影響
花青素具有抗腫瘤的生物特性,可抑制癌細胞的體外增殖、遷移和侵襲[9,10]。研究發現,從多種植物中提取的花青素在一定的時間和劑量范圍內均可抑制人肝癌HepG2細胞的增殖[11,12]。本試驗CCK-8結果顯示,濃度為25 μg/mL的黑果枸杞花青素作用于人肝癌HepG2細胞12、24和36 h時均可有效抑制人肝癌HepG2細胞的活力;濃度為50 μg/mL的黑枸杞花青素刺激人肝癌HepG2細胞24 h后,人肝癌HepG2細胞的活力也顯著降低;黑枸杞花青素作用24 h和36 h時,25 μg/mL黑果枸杞花青素較其他濃度組對人肝癌細胞的抑制作用更佳,其中,黑枸杞花青素作用24 h時,25 μg/mL黑果枸杞花青素對人肝癌HepG2細胞的抑制作用最為顯著,因此篩選出黑枸杞花青素最佳作用濃度和最佳作用時間分別為25 μg/mL和24 h。此外,EdU和細胞劃痕結果顯示,25 μg/mL黑果枸杞花青素誘導24 h后,可有效抑制人肝癌HepG2細胞增殖和遷移。
MOB1蛋白大型抑癌基因1/2(LATS1/2)激酶的關鍵調控因子,哺乳動物MST1-MOB 1復合物已被證明能抑制Yes相關蛋白(Yes-associated protein,YAP)的致癌活性,并參與多種腫瘤的發生[13]。YAP表達失調在人類癌癥中較為常見,YAP活性的降低可有效抑制癌細胞增殖和肝的過度生長[14]。LATS是公認的抑癌基因,在抑制癌細胞增殖的過程中可參與多條信號通路[15]。有研究表明,LATS缺失會引起惡性外周神經鞘膜瘤細胞的過度增殖[16]。LATS基因表達上調除可以抑制細胞的增殖外,還可有效抑制細胞的遷移[17]。研究發現哺乳動物LATS1和LATS2通過負調控不同的細胞周期檢查點來控制細胞的增殖,LATS2異位表達可使細胞停滯在G1/S期[18],而細胞在G1-S期的轉換主要由周期蛋白依賴性激酶4(CDK4)進行調控[19]。本研究結果顯示,用25 μg/mL的黑果枸杞花青素誘導人肝癌HepG2細胞24 h,增殖關鍵因子LATS1、LATS2和MOB1的mRNA表達被顯著上調,YAP的mRNA表達和CDK4的蛋白表達均被顯著下調。說明黑果枸杞花青素可通過抑制CDK4的表達使人肝癌HepG2細胞周期停滯在G1/S期并通過上調LATS1、LATS2、MOB1和下調YAP的mRNA表達來抑制人肝癌HepG2細胞的增殖。
3.3 黑果枸杞花青素對人肝癌HepG2細胞自噬作用的影響
近年來,細胞自噬在治療人類疾病方面的作用逐漸引起重視。當自噬發生時,Beclin-1被激活,進而活化LC3基因(包括LC3-I和LC3-II兩種亞型)中的LC3-I,而后LC3-I被修飾成LC3-II并通過融合蛋白轉位至自噬體膜,從而促進自噬體的成熟[20]。已有研究證實LC3-II的含量與自噬泡數量成正比,因此自噬程度與LC3-II 的表達水平呈正相關[21]。本研究發現,濃度為25 μg/mL 的黑果枸杞花青素人肝癌HepG2細胞24 h后可有效上調Beclin-1在mRNA水平的表達,并有效上調LC3-II在mRNA和蛋白水平的表達,說明黑果枸杞花青素可通過上調Beclin-1和LC3-II的表達進而誘導人肝癌HepG2細胞自噬。
研究發現,細胞自噬對肝癌的抑制作用可受到許多信號通路的調控,其中AMPK信號通路在激活細胞自噬的過程中起著重要作用[22]。Huang等[23]發現,增強肝癌細胞中的酮分解水平可有效抑制AMPK信號通路來抑制肝癌細胞發生過度自噬,從而促進肝癌細胞的增殖。本研究發現,濃度為25 μg/mL的黑果枸杞花青素作用24 h可明顯上調AMPK的mRNA表達和p-AMPK的蛋白表達,說明黑果枸杞花青素可能通過激活AMPK信號通路來誘導人肝癌HepG2細胞發生自噬進而抑制肝癌細胞的增殖,與Huang等研究結果一致。此外,mTOR信號通路在細胞自噬的過程中也扮演著重要的角色。Wang[24]和Jia等[25]發現,硫化氫和組蛋白乙酰轉移酶P300/CBP相關因子(P300/CBP-associa-ted factor,PCAF)均可通過抑制mTOR信號轉導途徑誘導肝癌細胞發生自噬,進而抑制人肝癌細胞HepG2細胞的增殖與遷移。本研究發現,濃度為25 μg/mL的黑果枸杞花青素作用24 h可有效下調p-mTOR的蛋白表達,說明黑果枸杞花青素可能通過抑制mTOR信號通路來促進人肝癌HepG2細胞發生自噬進而抑制人肝癌細胞HepG2細胞的增殖與遷移,與Wang和Jia等研究結果一致。
4 結論
綜上所述,通過HPLC-ESIQ-TOF分析從黑果枸杞花青素提取物中確認出5種花色苷,實現了復雜中藥成分快速分離分析,為黑果枸杞微量成分分析提供了一種有效分析方法。此外,結果發現25 μg/mL黑果枸杞花青素誘導24 h時可有效抑制人肝癌HepG2細胞的增殖和遷移,促進人肝癌HepG2細胞自噬,為黑果枸杞花青素的深度開發奠定了一定的理論基礎。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
