姜黃素增強(qiáng)PTEN野生型三陰性乳腺癌細(xì)胞對(duì)PARP1抑制劑敏感性的研究
2020-12-09 06:57:04孫偲豐楊彥琴玄基澤秦元華茅衛(wèi)鋒
中國(guó)比較醫(yī)學(xué)雜志 2020年11期
關(guān)鍵詞:乳腺癌
孫偲豐,王 瑩,楊彥琴,玄基澤,張 斌,秦元華,茅衛(wèi)鋒*
(1.大連醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院生物技術(shù)系,遼寧 大連 116044; 2.大連醫(yī)科大學(xué)附屬第二醫(yī)院腫瘤放射治療科,遼寧 大連 116011; 3.大連醫(yī)科大學(xué)附屬第二醫(yī)院病理科,遼寧 大連 116011; 4.大連醫(yī)科大學(xué)第一附屬醫(yī)院腫瘤科,遼寧 大連 116011; 5.大連醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院寄生蟲(chóng)系,遼寧 大連 116044)
乳腺癌是發(fā)生于女性中最?lèi)盒缘哪[瘤之一,每年約有458,000人死于乳腺癌[1-2]。三陰性乳腺癌是雌激素受體(ER)、孕激素受體(PR)和原癌基因Her-2都為陰性的一種乳腺癌。由于是三陰性,常規(guī)靶向藥物無(wú)法對(duì)其產(chǎn)生作用,只能按照常規(guī)放化療治療,但這種治療效果較差,伴隨低療效、高復(fù)發(fā)的特性。PARP1抑制劑作為一種新型的靶向PARP1的分子靶向藥物,近年來(lái)在乳腺癌治療中表現(xiàn)出較好的療效。PARP1是PARP家族的主要成員,其可將ADP-核糖基團(tuán)轉(zhuǎn)移至靶蛋白[3-5]。一旦DNA斷裂發(fā)生,PARP1作為單鏈斷裂(SSB)修復(fù)中的關(guān)鍵因子可以感知損傷并快速結(jié)合到斷裂位點(diǎn)并使自身核糖基化,并進(jìn)一步催化靶蛋白的核糖基化,招募DNA修復(fù)蛋白進(jìn)行DNA修復(fù)[6]。因此PARP1抑制劑奧拉帕尼作為抗腫瘤藥物具有良好效果,并已在臨床上用于治療乳腺癌和卵巢癌[7-8]。據(jù)報(bào)道,奧拉帕尼可有效抑制BRCA1和PTEN缺陷腫瘤細(xì)胞的生長(zhǎng),顯示出PARP1和BRCA1/PTEN的合成致死性。然而,野生型PTEN的腫瘤細(xì)胞表現(xiàn)出對(duì)PARP1抑制劑的不敏感性[9],因此,急需要尋找可以增加野生型PTEN腫瘤細(xì)胞對(duì)奧拉帕尼敏感性的協(xié)同劑。
磷酸酶和張力蛋白同源物(PTEN)是位于人類(lèi)基因組10q23.3上的抑癌基因表達(dá)的蛋白,PTEN通過(guò)參與PI3K途徑而在細(xì)胞增殖中起作用[10-11]。PTEN突變經(jīng)常發(fā)生在各種類(lèi)型的癌癥中[12-13]。PTEN還促進(jìn)DNA修復(fù)以維持染色體的穩(wěn)定性[14-15]。PTEN能增強(qiáng)腫瘤細(xì)胞對(duì)化療藥物抗性[16]。PTEN與復(fù)制蛋白A1協(xié)同作用,可以保護(hù)DNA復(fù)制叉并增強(qiáng)RAD51介導(dǎo)的同源重組(HR)。然而,PTEN在DNA修復(fù)中的具體作用仍未明確[9,15]。
姜黃素是一種從姜黃根中分離出來(lái)的天然化合物,具有抗氧化,抗炎和抗腫瘤活性[17-22]。姜黃素易獲得,價(jià)格便宜,常用作膳食調(diào)味品[23-24]。據(jù)報(bào)道,姜黃素可抑制不同類(lèi)型腫瘤細(xì)胞的生長(zhǎng),并使癌細(xì)胞對(duì)阿霉素和5-FU敏感[25]。本研究主要探討了姜黃素對(duì)PTEN介導(dǎo)的DNA修復(fù)的作用及其潛在機(jī)制。這些結(jié)果可擴(kuò)展姜黃素在PTEN野生型三陰性乳腺癌治療中的應(yīng)用,也可以為異常PTEN腫瘤細(xì)胞的個(gè)性化治療提供理論基礎(chǔ)。
1 材料和方法
1.1 實(shí)驗(yàn)細(xì)胞
人乳腺癌細(xì)胞系BT549從美國(guó)組織培養(yǎng)物收集中心(ATCC,美國(guó))購(gòu)得,是PTEN缺陷的三陰性乳腺癌細(xì)胞系。細(xì)胞在含有10% FBS(Biological Industries,Kibbutz Beit Haemek,以色列),1%青霉素和鏈霉素(Gibico,美國(guó))的RPMI 1640(Hyclone,美國(guó))中培養(yǎng)。本實(shí)驗(yàn)室穩(wěn)轉(zhuǎn)PTEN基因到BT549中,獲得含PTEN基因的BT549,代表野生型BT549。
1.2 主要試劑
PTEN的抗體(sc-9145)購(gòu)自Santa Cruz(美國(guó));β-肌動(dòng)蛋白(66009-1-1g)、PARP1(13371-1-AP)、微管蛋白(11224-1AP)、RAD51(14961-1-AP)的抗體購(gòu)自Proteintech(美國(guó))。PARP抑制劑奧拉帕尼(S1060)、姜黃素(S1814)購(gòu)自Selleck(美國(guó));苯甲基磺酰氟(PMSF);蛋白酶抑制劑(B14001,Selleck,美國(guó));磷酸酶抑制劑(B15001,Selleck,美國(guó));Fluorometer Qubit 2.0(Invitrogen,美國(guó));MTT、NaCl、EDTA、Tris、肌氨酸鈉、Trixton X-100、DMSO、NaOH(生工,中國(guó))。
1.3 實(shí)驗(yàn)方法
1.3.1 蛋白質(zhì)印跡
用含1 mmol/L苯甲基磺酰氟(PMSF),1×蛋白酶抑制劑和1×磷酸酶抑制劑的冰冷RIPA緩沖液裂解細(xì)胞;Fluorometer Qubit 2.0測(cè)定總蛋白濃度;使用10%SDS-PAGE凝膠分離細(xì)胞裂解物;將凝膠上的蛋白轉(zhuǎn)移至PVDF膜,使用各種抗體(稀釋比1∶500)檢測(cè)蛋白質(zhì)。
1.3.2 MTT檢測(cè)
將細(xì)胞(4000個(gè)細(xì)胞/孔)接種在96孔板中。待細(xì)胞貼壁后,加入藥物,與細(xì)胞一起溫育48 h。取出藥物,然后將3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物(MTT)加入96孔板,與細(xì)胞一起溫育4 h。除去培養(yǎng)基后加入150 μL的DMSO,492 nm處讀取吸光度。

注:A:通過(guò)蛋白質(zhì)印跡法分別檢測(cè)轉(zhuǎn)染PTEN基因、空載體的BT549細(xì)胞中PTEN的表達(dá);B:PTEN細(xì)胞和EV(空載體)細(xì)胞通過(guò)MTT法測(cè)定的生長(zhǎng)曲線。PTEN:穩(wěn)定轉(zhuǎn)染PTEN的BT529細(xì)胞,EV:轉(zhuǎn)染空載體的BT549細(xì)胞。圖1 PTEN基因的BT529細(xì)胞和轉(zhuǎn)染空載體(EV)的BT549細(xì)胞的生長(zhǎng)曲線Note. A, Expression of PTEN in BT549 cells transfected with PTEN gene and empty vector was detected by Western blot. B, Growth curves of PTEN cells and EV cells were tested by MTT assay. PTEN, BT529 cells stably transfected with PTEN. EV, BT549 cells transfected with empty vector.Figure 1 Growth curves of PTEN-transfected BT529 cells and empty vector-transfected BT549 cells
1.3.3 彗實(shí)實(shí)驗(yàn)
40 μmol/L姜黃素或80 μmol/L奧拉帕尼處理細(xì)胞后,取5000個(gè)細(xì)胞包埋在低熔點(diǎn)瓊脂糖的載玻片上。將載玻片浸入冰冷的裂解緩沖液(2.5 mol/L NaCl,100 mmol/L EDTA,10 mmol/L Tris,1%肌氨酸鈉,1% Trixton X-100,10%DMSO)裂解40 min。然后將載玻片放入堿性緩沖液(1 mmol/L EDTA,300 mmol/L NaOH)中電泳。用溴化乙錠染色后,顯微鏡分析結(jié)果。
1.4 統(tǒng)計(jì)學(xué)方法
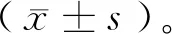
2 結(jié)果
2.1 PTEN上調(diào)RAD51的表達(dá)增強(qiáng)DNA修復(fù)
如圖1所示,野生型BT549與BT549的細(xì)胞存活曲線(圖1A)顯示兩者間沒(méi)有生長(zhǎng)差異(圖1B)。
MTT測(cè)定顯示EV細(xì)胞對(duì)奧拉帕尼敏感,而PTEN細(xì)胞對(duì)奧拉帕尼耐藥。這些結(jié)果表明野生型PTEN細(xì)胞對(duì)新型分子靶向藥物奧拉帕尼耐受(圖2A)。隨后,蛋白質(zhì)印跡實(shí)驗(yàn)檢測(cè)了兩種細(xì)胞RAD51的水平(圖2B),顯示PTEN野生型細(xì)胞的RAD51表達(dá)更高,表明PTEN增加RAD51介導(dǎo)的同源重組修復(fù)以促進(jìn)DNA修復(fù)。
2.2 姜黃素增強(qiáng)奧拉帕尼誘導(dǎo)的DNA斷裂
接下來(lái)分析了姜黃素對(duì)PTEN促進(jìn)DNA修復(fù)的影響。用奧拉帕尼處理PTEN野生型細(xì)胞24 h后,通過(guò)彗星試驗(yàn)檢測(cè)DNA斷裂。圖中顯示姜黃素誘導(dǎo)DNA的斷裂。更重要的是,與單種藥物產(chǎn)生的彗尾相比,姜黃素與奧拉帕尼的聯(lián)合使用明顯增加了彗尾(圖3A、3B)。這些結(jié)果表明姜黃素促進(jìn)PTEN野生型細(xì)胞中奧拉帕尼誘導(dǎo)的DNA斷裂。
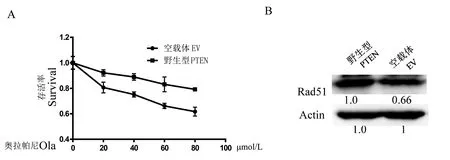
注:A:用不同劑量的奧拉帕尼處理24 h后,與轉(zhuǎn)染空載體的BT549細(xì)胞相比,PTEN細(xì)胞具有olaprib抗性;B:蛋白質(zhì)印跡顯示,與EV細(xì)胞中的RAD51水平相比,PTEN上調(diào)RAD51的表達(dá)。圖2 PTEN上調(diào)RAD51表達(dá),增強(qiáng)PTEN野生型BT549對(duì)PARP1抑制劑奧拉帕尼的耐受Note. A, After treatment with different doses of Olaparib for 24 hours, PTEN cells were resistant to olaprib compared to BT549 cells transfected with an empty vector. B, Western blot shows that PTEN up-regulates RAD51 expression compared to RAD51 levels in EV cells.Figure 2 PTEN upregulates RAD51-mediated resistance of PTEN wild-type BT549 to the PARP1 inhibitor, Olaparib

注: A:用奧拉帕尼或姜黃素處理轉(zhuǎn)染PTEN的BT549細(xì)胞12 h。彗星實(shí)驗(yàn)顯示,40 μmol/L姜黃素增加了彗尾,并促進(jìn)了奧拉帕尼(80 μmol/L)誘導(dǎo)的PTEN細(xì)胞的彗尾;B:分析彗星實(shí)驗(yàn)中的30個(gè)彗尾。單因素方差分析和 Turkey’s檢驗(yàn),與奧拉帕尼治療組相比,***P <0.001。圖3 姜黃素促進(jìn)奧拉帕尼處理后的PTEN細(xì)胞中的DNA斷裂Note. A, BT549 cells transfected with PTEN were treated with Olaparib or curcumin for 12 hours. The comet assay showed that 40 μmol/L curcumin increased the comet tail and promoted the comet tail of PTEN cells induced by Olaparib (80 μmol/L). B, Analysis of 30 comet tails in comet experiments. One-way analysis of variance and Turkey’s test, compared with the Olaparib treatment group,***P<0.001.Figure 3 Curcumin promotes DNA breaks in PTEN cells treated with Olaparib
2.3 姜黃素降低PTEN乳腺癌細(xì)胞中RAD51的表達(dá)
隨后分析了姜黃素增強(qiáng)PTEN野生型乳腺癌細(xì)胞對(duì)奧拉帕尼敏感性的潛在機(jī)制。用姜黃素處理后,RAD51水平以劑量依賴(lài)性方式降低(圖4)。因此,姜黃素會(huì)降低RAD51的表達(dá),從而減輕PTEN介導(dǎo)的DNA修復(fù)。此外,我們還發(fā)現(xiàn)姜黃素降低了PARP1的水平,這表明姜黃素可能同樣影響PARP1參與的DNA修復(fù)(圖4)。

注:蛋白質(zhì)印跡結(jié)果顯示姜黃素降低了PTEN細(xì)胞中Rad51和PARP1的水平。每個(gè)Western數(shù)值均由三次實(shí)驗(yàn)完成,其中對(duì)照數(shù)值設(shè)定為1。Turkey’s檢驗(yàn),***P<0.001。圖4 姜黃素降低了PTEN細(xì)胞中RAD51的水平Note. Western blot results showed that curcumin reduced the levels of RAD51 and PARP1 in PTEN cells. Each Western value was completed by three experiments. The control value was set 1. Turkey’s test,***P<0.001.Figure 4 Curcumin decreases the levels of RAD51 which is up-regulated in PTEN cells
2.4 姜黃素使PTEN野生型細(xì)胞對(duì)奧拉帕尼敏感
我們用MTT實(shí)驗(yàn)檢測(cè)了姜黃素和奧拉帕尼聯(lián)合處理的PTEN野生型細(xì)胞的生長(zhǎng)曲線,與單藥奧拉帕尼或姜黃素相比,顯示出姜黃素以劑量依賴(lài)性方式使PTEN野生型細(xì)胞對(duì)奧拉帕尼敏感(圖5A、5B)。這些結(jié)果表明,姜黃素與奧拉帕尼聯(lián)合使用時(shí),PTEN野生型癌細(xì)胞更加敏感。

注: A:姜黃素單藥處理PTEN細(xì)胞與EV細(xì)胞后的存活曲線;B:MTT實(shí)驗(yàn)檢測(cè)姜黃素與奧拉帕尼聯(lián)合使用后,PTEN細(xì)胞的存活曲線。單因素方差分析和Turkey’s檢驗(yàn),與奧拉帕尼治療組相比,***P<0.001,**P <0.01,*P <0.05。圖5 姜黃素使轉(zhuǎn)染PTEN的BT549細(xì)胞對(duì)奧拉帕尼敏感Note. A, Survival curves of curcumin monotherapy on PTEN cells and EV cells. B, MTT test to detect the survival curve of PTEN cells after curcumin was combined with Olaparib. One-way analysis of variance and Turkey’s test, compared with Olaparib treatment group,***P <0.001,**P <0.01,*P <0.05.Figure 5 Curcumin sensitizes PTEN-transfected BT549 cells to Olaparib
3 討論
PTEN是癌細(xì)胞中高度突變的抑癌基因,是癌癥治療的潛在靶標(biāo)靶標(biāo)[12]。最近的報(bào)道顯示PTEN具有DNA修復(fù)能力,使得PTEN野生型腫瘤細(xì)胞具有DNA損傷藥物抗性[9,16], 因此研究PTEN引起的腫瘤化療耐藥性具有重要意義。奧拉帕尼是PARP1抑制劑,通過(guò)抑制PARP1活性以抑制單鏈斷裂修復(fù)[7]。PARP1抑制劑不直接誘導(dǎo)DNA斷裂,因此它具有低毒性并且是有前景的抗腫瘤藥物。奧拉帕尼已被臨床用于治療卵巢癌和乳腺癌。然而,奧拉帕尼的作用依賴(lài)于腫瘤細(xì)胞的遺傳背景。BRCA異常癌細(xì)胞對(duì)奧拉帕尼敏感,而B(niǎo)RCA和PTEN野生型細(xì)胞對(duì)奧拉帕尼耐藥[7]。由于PTEN在DNA修復(fù)中的功能尚不清楚,因此很少報(bào)道能使PTEN野生型腫瘤細(xì)胞敏感的藥物。據(jù)報(bào)道RI-1是一種RAD51抑制劑,可增加PTEN腫瘤細(xì)胞對(duì)順鉑的敏感性。然而,RI-1也不是臨床試劑。此外,抑制RAD51并不能完全降低PTEN的DNA修復(fù)能力。
幾個(gè)世紀(jì)以來(lái),姜黃素一直被用作食用性香料,且在I期人體試驗(yàn)中,受試者每天使用量高達(dá)8000 mg,持續(xù)3個(gè)月仍沒(méi)有顯示出毒性[23]。據(jù)報(bào)道,姜黃素還具有抗腫瘤活性并可抑制多種DNA修復(fù)途徑[25],姜黃素干預(yù)N-甲基亞硝基脲誘發(fā)膀胱癌[26]。針對(duì)三陰性乳腺癌的耐化療藥物特性[27],本研究分析了姜黃素對(duì)于PTEN的DNA修復(fù)能力的影響,同時(shí)分析了姜黃素對(duì)奧拉帕尼處理PTEN野生型三陰性乳腺癌細(xì)胞敏感性的影響。本研究發(fā)現(xiàn)PTEN促進(jìn)奧拉帕尼誘導(dǎo)DNA損傷的修復(fù),細(xì)胞生長(zhǎng)曲線顯示PTEN有助于腫瘤細(xì)胞抵抗奧拉帕尼。用姜黃素處理后,彗星實(shí)驗(yàn)顯示PTEN野生型腫瘤細(xì)胞中的DNA斷裂增加。MTT實(shí)驗(yàn)顯示姜黃素增加PTEN野生型腫瘤細(xì)胞對(duì)奧拉帕尼的敏感。我們的研究結(jié)果表明,姜黃素可以抑制PTEN促進(jìn)的DNA修復(fù)。
RAD51是HR中的關(guān)鍵因子,據(jù)報(bào)道,它在PTEN促進(jìn)DNA的修復(fù)中起較為重要的作用[9]。我們發(fā)現(xiàn)姜黃素明顯降低了RAD51的水平。該結(jié)果表明姜黃素干擾RAD51介導(dǎo)的HR并且可以通過(guò)降低RAD51水平影響PTEN介導(dǎo)的DNA修復(fù)。另外我們還發(fā)現(xiàn)姜黃素明顯降低了PARP1的水平。PARP1是修飾蛋白質(zhì)的聚-ADP-核糖基化(PARylation)的酶,可添加的多聚ADP-核糖(PAR)至靶標(biāo)蛋白進(jìn)而改變蛋白的電荷從而影響它們與DNA的相互作用[5]. 許多DNA修復(fù)因子,如XRCC1,Ku70和DNA-PKcs均是PARP1的靶標(biāo)。由于PARP1影響DNA修復(fù)因子和DNA斷裂位點(diǎn)之間的相互作用,因此PARP1被認(rèn)為參與多種DNA修復(fù)途徑。姜黃素降低PARP1的水平,表明姜黃素除了對(duì)RAD51介導(dǎo)的HR有影響外還可以干擾其他DNA修復(fù)途徑。總之,本文的研究表明姜黃素可以抑制PTEN的DNA修復(fù)并使PTEN野生型癌細(xì)胞對(duì)奧拉帕尼敏感。
綜上所述,這些結(jié)果表明姜黃素通過(guò)抑制RAD51介導(dǎo)的同源重組達(dá)到抑制PTEN的DNA修復(fù)能力,從而使PTEN野生型乳腺癌細(xì)胞對(duì)PARP1抑制劑敏感。這些發(fā)現(xiàn)將促進(jìn)姜黃素在具有不同基因型PTEN腫瘤細(xì)胞的臨床個(gè)體化治療中的應(yīng)用。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
現(xiàn)代臨床醫(yī)學(xué)(2022年1期)2022-02-12 02:04:58
甘肅科技(2020年20期)2020-04-13 00:30:42
中國(guó)生殖健康(2019年2期)2019-08-23 08:11:42
中國(guó)生殖健康(2019年6期)2019-01-06 09:20:12
中國(guó)生殖健康(2019年5期)2019-01-06 09:16:40
幸福家庭(2019年14期)2019-01-06 09:15:38
祝您健康(2018年5期)2018-05-16 17:10:16
癌癥進(jìn)展(2016年9期)2016-08-22 11:33:20
中國(guó)組織化學(xué)與細(xì)胞化學(xué)雜志(2016年4期)2016-02-27 11:16:08
