濱海鹽沼濕地植被凈初級生產力變化對土壤因子的響應
2020-11-14 07:07:34崔林林李國勝歐陽寧雷陳吉龍廖華軍趙耕樂
生態學報 2020年19期
崔林林,李國勝,歐陽寧雷,陳吉龍,廖華軍,趙耕樂
1 成都信息工程大學資源環境學院,成都 610225 2 中國科學院地理科學與資源研究所陸地表層格局與模擬院重點實驗室,北京 100101 3 中國地質調查局濱海濕地生物地質重點實驗室,青島 266071 4 中國科學院重慶綠色智能技術研究院, 重慶 400714
植被凈初級生產力(Net Primary Productivity, NPP)是指植物通過光合作用在單位面積、單位時間所同化的碳總量中扣除自養呼吸后的剩余部分,它是生態系統中其他生物生存和繁衍的物質基礎,是碳循環的重要環節,直接反映植物固碳的效率[1]。與農業生態系統、草地生態系統和森林生態系統(包括熱帶雨林生態系統)相比,濱海鹽沼濕地具有更高的生產力,在全球碳循環中具有重要作用[2]。濱海鹽沼濕地NPP是植物生命活動的結果,受到植物種類、氣候、水文、土壤性質等因子的影響,并且這些因子之間也相互作用、相互制約,共同決定植被NPP的大小[3-5]。因此,研究這些因子對濱海鹽沼濕地NPP的影響對于理解它們之間相互作用機理,更好地進行濕地管理和保護具有重要意義。
氣候因子(如太陽輻射、CO2濃度、氣溫等)通過影響植被的光合作用效率和物候、土壤養分的有效性和微生物活動等來影響植被NPP[6-10]。對于一個空間尺度相對較小的區域,在氣候變化尺度上,長期氣候變化是決定植被NPP長期演變的主導因素之一;在年變化尺度上,溫度、水分等氣候要素的年內變化,雖然影響植被NPP的逐月分布狀態,但氣候因子可能由于其空間差異性較小,而不是導致植被NPP空間分異的主導因素。植被類型和土壤因子可能是較小空間尺度植被NPP空間分異的決定因素,而同種植被類型內部NPP空間分異的主導因素可能是土壤因子。
濱海鹽沼濕地是陸地生態系統的一種重要類型,也是受營養限制的生態系統[11]。營養限制的研究由單營養限制向多營養聯合限制發展[12]。氮(N)被認為是影響陸地生態系統植被生產力的關鍵因素[13-15]。目前,已有許多研究對生態系統中N、磷(P)及其組合對植被生長的限制作用進行了研究,結果表明植被生產力受N和P限制,且兩者具有協同作用[16-17]。Fay等通過人為控制實驗研究了N、P、鉀(K)及其組合對植被生產力的影響,除佐證了N、P的協同作用外,還指出了K和微量元素限制的重要性[18]。在草地或其他植被生長中,追施營養也受到了越來越多的關注[19]。無論如何,濱海鹽沼濕地植被生產力的營養多重限制,特別是在不同水鹽條件下營養物質的共同限制潛力的研究還較少[12,18,20],這也就意味著需要進一步研究濱海鹽沼濕地植被NPP的營養限制大小和程度。
水鹽是濱海鹽沼濕地形成的重要因子,影響著植被分布格局,造成植被NPP的空間差異。很多研究表明濱海鹽沼濕地植被分布受土壤水分和鹽度的雙重影響,有各自的生態閾值[21]。土壤水鹽的變化能夠改變植被群落的分布格局、植被的形態結構來影響生產力:濕地水鹽梯度的變化決定著濕地植被群落的分布格局[22-23];水位深度的變化能夠改變濕地植物的形態結構來影響生產力[24]。但目前,水鹽對植被生產力的影響還缺乏定量的認識,需要進一步的深入研究。
綜上所述,對于空間尺度較小的區域,影響濱海鹽沼濕地植被NPP空間分異的因子主要是土壤因子。這些因子單獨或以不同組合形式對濱海鹽沼濕地植被NPP的影響大小、程度及其組合的限制形式都還不甚清楚。因此,本文以遼河三角洲濱海鹽沼濕地為研究對象,采用遙感和地面調查相結合方法,研究土壤因子對鹽沼濕地植被NPP空間分異的影響。該研究結果將有助于濕地植被的保護和修復,以期為該領域的進一步研究提供參考。
1 材料和方法
1.1 研究區介紹
遼河三角洲濱海濕地位于遼寧省西南部遼河平原南端,是我國重要的河口濕地之一,由來自遼河、雙臺子河、大凌河、小凌河和大清河等沖積而成,行政區劃上包括盤錦市和營口市全部區域,及錦州部分地區。本文研究區如圖1所示,位于12110′—12230′ E, 北緯4030′ —4130′ N之間,面積約6000 km2。該區域地貌屬于沖海積平原,地勢平坦,氣候為暖溫帶大陸性半濕潤季風氣候,四季分明、雨熱同期:年平均溫度8.6 ℃、年平均降雨量約為612 mm、年平均蒸發量為1670 mm。受海洋、河流和陸地交互、淡咸水交互的影響,研究區發育了以蘆葦(Phragmitesaustralis)、翅堿蓬(Suaedapterantha)、檉柳(TamarixchinensisLour.)、獐毛(Aeluropussinensis)、香蒲(TyphaorientalisPresl)等作為建群種的濕地植物群落,其中蘆葦沼澤面積達到756 km2,是該區域分布最廣、面積最大的植物群落[21],也是亞洲第一大蘆葦濕地[25]。

圖1 研究區示意圖Fig.1 Location and extent of research area
1.2 數據收集及預處理
1.2.1遙感數據
本文所用遙感數據包括2011年的MODIS(Moderate-resolution Imaging Spectroradiometer)數據、2011年6月19日和2009年7月15日的Landsat-5 TM數據、DEM(Digital Elevation Model)數據,具體數據選擇如表1所示。MODIS數據利用MRT(MODIS Reprojection Tool)軟件完成重投影、格式轉換和重采樣等預處理,制成分辨率為1000 m、投影坐標系為橫軸墨卡托投影,橢球體為WGS84的MODIS數據集;基于TM的4級數據和DEM數據,對于TM的1級數據進行的預處理工作包括幾何校正、輻射定標、大氣校正、圖像鑲嵌和剪切。其中,大氣校正是定量遙感研究的關鍵步驟,研究表明改進的基于圖像的暗目標減法能較好地應用于海岸帶遙感數據[26],本研究選用該方法對遙感影像進行大氣校正。
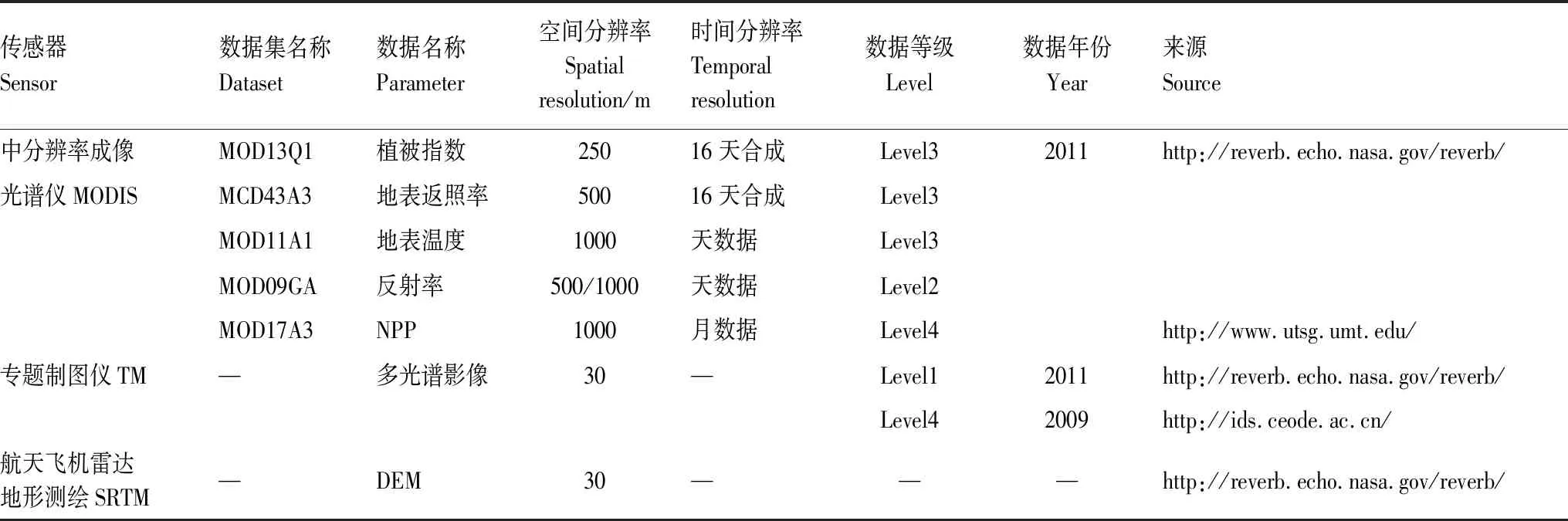
表1 本研究所用遙感數據Table 1 Remote sensing data used in this study
1.2.2氣象數據
研究區內有3個氣象站:錦州、大洼和營口,數據來自中國氣象科學數據共享服務網(http://cdc.cma.gov.cn/)。數據包括日照時數、風速、相對濕度、氣溫、蒸散發和太陽輻射數據等。太陽總輻射用?ngstr?m-Prescott模型來估算,其他氣象數據利用高度訂正的反距離權重法進行空間插值。
1.2.3土地利用數據
土地利用數據是基于2011年的Landsat-5 TM數據,利用目視解譯方法提取,共分為20類(圖2)。景觀類型主要包括天然濕地(淺海水域、淺水沙洲、灘涂、河流、灘地、草甸、葦田)、人工濕地(水田、水庫、水池、鹽池、養殖水體)、非濕地(旱地、林地、灌叢、城鄉居民點、工礦、道路、防護堤和裸地)。

圖2 2011年土地利用圖Fig.2 Land use map in 2011
1.2.4野外調查和實驗室分析數據
2011年10月進行了野外調查。由于濕地可進入條件的限制,野外生物量測量樣點基本沿道路布設,共設樣點30個(15個用于模擬最大光能轉化率,另外15個用于模型驗證)。野外生物量調查采用五點交叉采樣法:大樣地面積設置為30 m×30 m,每個樣地的四角和中心各設置一個2m×2 m小樣方,按收獲法進行生物量調查。測定樣方地上總鮮重包括莖、葉和凋落物,每一樣方取300—500 g左右的樣品,用于測定干物質比率。樣品帶回實驗室利用烘箱在80 ℃下烘至恒重,計算樣品的干物質比率,再根據樣方鮮重計算樣方內地上生物量總干重,利用同一樣地5個樣方的均值代表樣地單位面積地上生物量。由于蘆葦的根冠比較大,所以本研究采用邵成等[27]的研究結論和野外采樣點根冠比的均值(1.722)計算樣地單位面積總生物量。生物量轉換為碳含量還需要“碳轉換系數”,研究表明木質、草和葉成分的碳轉換系數分別為0.5、0.45和0.45[28-29]。由于蘆葦是草本植物,所以設定轉換系數為0.45。
研究區濕地鹽度呈明顯的水平變化規律,由內陸向濱海地區遞增[30]。在對研究區考查的基礎上,結合土地利用圖、NPP空間分布和水鹽梯度情況,在蘆葦沼澤區域,大致與河口海岸垂直的方向上,從濱海向內陸設置了52個土壤采樣點(圖 3)。利用土壤取樣器,鉆取50 cm深的土壤樣品,按10 cm間隔取樣,利用梅特勒-托利多國際股份有限公司生產的便攜式PH計和英國Delta-T公司生產的W.E.T土壤三參數儀現場獲得土壤的PH值、溫度(ST)、含水量(SW)、溶氧量(SO)和電導率(體積電導率ECb、孔隙電導率ECp)。
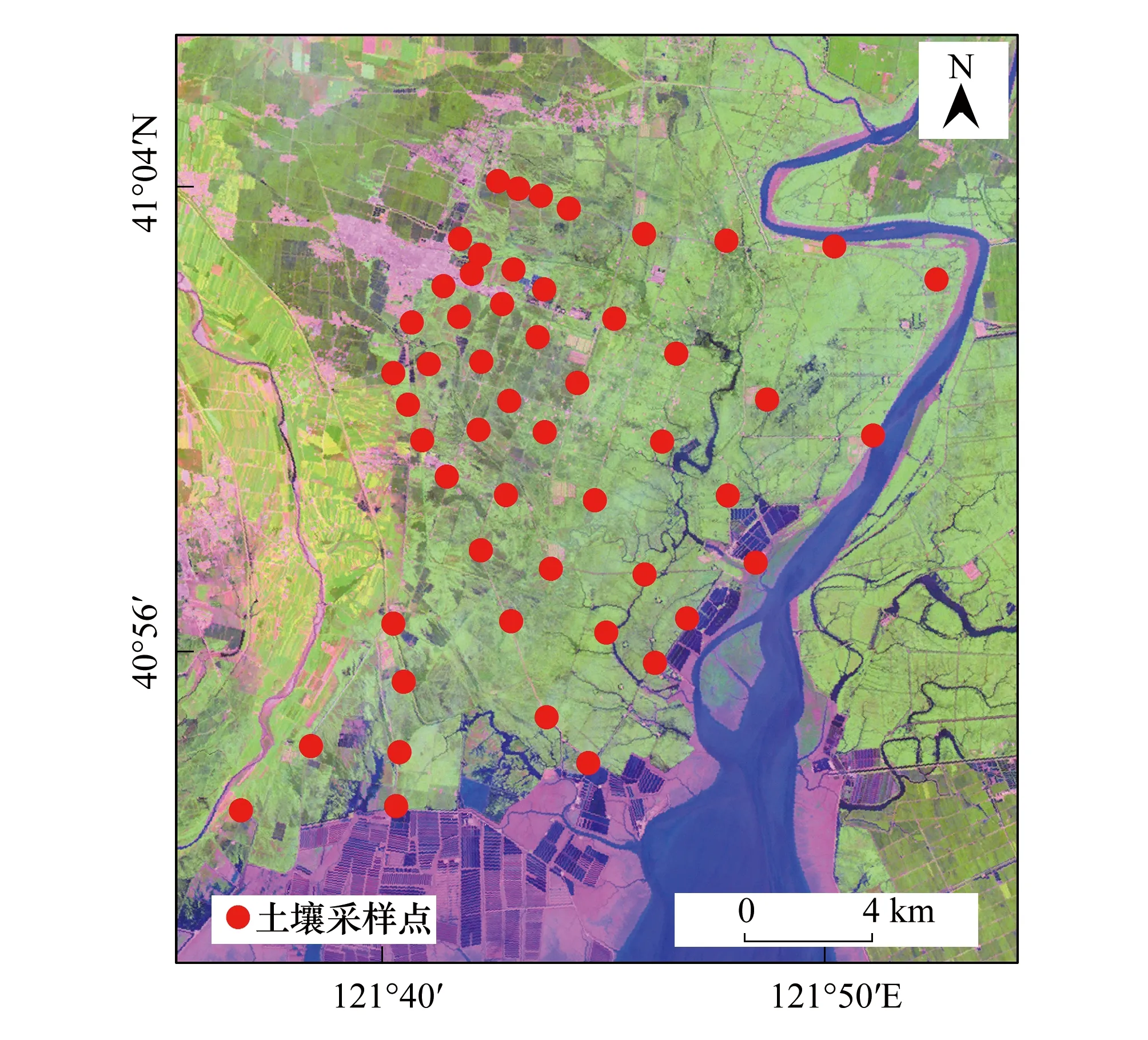
圖3 土壤樣點設置Fig.3 Setting of soil sampling points
土壤樣品被放入塑料盒,置于室內風干,用鑷子剔除動植物殘體和其他雜質(石塊等)。然后,利用球磨儀將風干土樣研細,過100目篩子,將樣品充分混合用于全碳(TC)、全氮(TN)、碳氮比(CN)、速效磷(AP)和速效鉀(AK)等理化性質測定。利用德國Elementar Vario Max CN碳氮元素分析儀測定的全碳、全氮、碳氮比;利用美國Perkin Elmer公司的Optima 5300DV電感耦合等離子體—原子發射光譜儀測定的速效磷和速效鉀。
2 研究方法
2.1 生產力估算方法
CASA(Carnegie-Ames-Stanford approach)模型是Potter于1993年提出的基于光能利用率的生產力估算模型[31]。原始的CASA模型主要采用遙感數據、氣象數據和土壤質地數據進行生產力反演,其表達式如下:
NPP=PAR×FPAR×ε
(1)
ε=Tg1×Tg2×Wg×εmax
(2)
式中,NPP為凈初級生產力,PAR為入射的光合有效輻射,FPAR為植被冠層對光合有效輻射的吸收分量,ε為實際光能利用率,Tg1和Tg2為溫度脅迫系數,Wg為水分脅迫系數,εmax為最大光能利用率。本研究利用SEBAL(Surface Energy Balance Algorithm for Land)模型[32]和FAO56-PM公式代替土壤水分子模型和Thornthwaite方法計算土壤水分脅迫系數。
2.2 因子敏感性分析方法
偏最小二乘方法是1983年由S. Wold和C. Albano等人提出的一種具有廣泛適用性的多元統計分析方法[33]。在偏最小二乘分析中變量投影重要性(variable importance in projection, VIP)指標可以說明變量X在解釋Y時作用的重要性,公式如下:
(3)
式中,VIPj是變量xj對解釋Y的重要性;whj是軸wh的第j個分量,被用于測量xj對構造th成分的邊際貢獻。基本思想是xj對Y的解釋是通過th來傳遞的,如果th對Y的解釋能力很強,而xj在構造th時又起到了重要的作用,則xj對Y的解釋能力就被視為很大。
2.3 影響因素分析方法
(1)本文擬通過把NPP和土壤因子的空間分布進行差異化分組,分析土壤因子對NPP空間分異的影響。因此,需要先對樣點數據進行空間化處理。經過試驗發現BP神經網絡隱含層和輸出層分別設為tansig、logsig和purelin,隱含層節點數設為3—30,輸入數據包括植被指數、距河流的距離、距海水的距離和高程數據,對土壤理化性質進行空間化結果較好。
(2)為了分析土壤因子對NPP空間分異的影響,根據灰度直方圖,將NPP數據和土壤因子空間化數據分成10個組;基于分組數據,將土壤全氮、速效磷和速效鉀的第一類(含量最低)賦予1,其他類賦予2,將賦值后的圖層進行疊加處理,各分組如表2所示。營養限制的存在和大小不僅和氣候、土壤發育或肥力有關,還與其他一個或多個營養限制有關,且NPP的多營養限制形式稱為聯合限制,包括協同限制、可加性限制和次可加性限制[18]。協同限制——多營養限制大于單營養限制之和;可加性限制——多營養限制等于單營養限制之和;次可加性限制——多營養限制小于單營養限制之和。

表2 分組結果Table 2 The grouped result
(3)利用處理響應比的自然對數計算各氮、磷、鉀處理影響的大小,公式如下[18]:
LRR=ln(NPP處理組/NPP對照組)
(4)
3 結果分析
3.1 NPP反演結果分析
由圖4可知,估計值和實測值的相關性較好,相關系數為0.89,擬合優度R2為0.78(P<0.01)。這說明CASA模型能被用于本研究區的NPP反演。
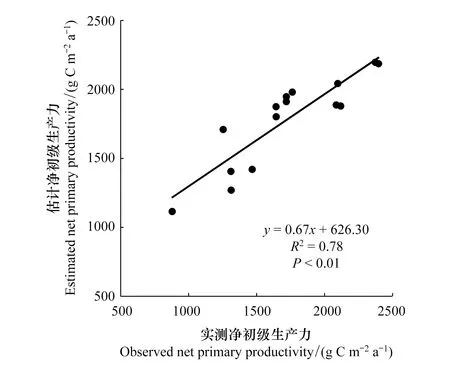
圖4 實測NPP與模型估計NPP的比較Fig.4 Comparison of the estimated and observed NPP
圖5是2011年研究區NPP的空間分布圖。由圖2和圖5可知,研究區NPP的取值范圍為31.66—2417.22 gCm-2a-1,且變異系數為43.71%,空間分異明顯;NPP空間分布大致呈現出以濕地核心區蘆葦沼澤為中心向外圍降低的環形空間分布格局,且區域NPP和植被分布格局具有較好的一致性。生產力最高的是中部的蘆葦沼澤,然后是外圍的水田和旱地,最后是林地和草甸。不同植被景觀交界處NPP呈階梯式急劇下降,空白區為非植被景觀點綴其中。這說明區域尺度上植被類型分布整體上決定了區域NPP的空間分異,但同種植被類型內部也具有較為明顯的區域分異特征,如蘆葦沼澤、水田和旱地的NPP變異系數分別為24.06%、15.08%和18.65%。因此,本文就以蘆葦沼澤為例,研究土壤因子對蘆葦沼澤NPP空間分異的影響。

圖5 2011年NPP空間分布圖Fig.5 Spatial variation of net primary productivity
3.2 NPP空間分異的影響因素
3.2.1因子敏感性分析
以蘆葦沼澤為例,將NPP作為自變量,土壤因子(TC、TN、AP、AK、CN、PH、SO、ST、SW、ECb、ECp)和DEM數據作為因變量構建偏最小二乘回歸模型,獲得變量投影重要性數值(表3)。由表3可知,土壤因子和地形因子對蘆葦沼澤NPP的敏感性從大到小排序為:AK>TC>TN>AP>ST>DEM>SW>CN>ECb>SO>PH>ECp。由表4可知,各因子間存在很強的相關性。為簡化分析過程和闡述的簡潔性,依據敏感性分析結果和各因子之間的相關性,本文選擇ECb、SW、TN、AP和AK等5個敏感性因子,分析各因子及其組合對濕地生態系統生產力的影響。
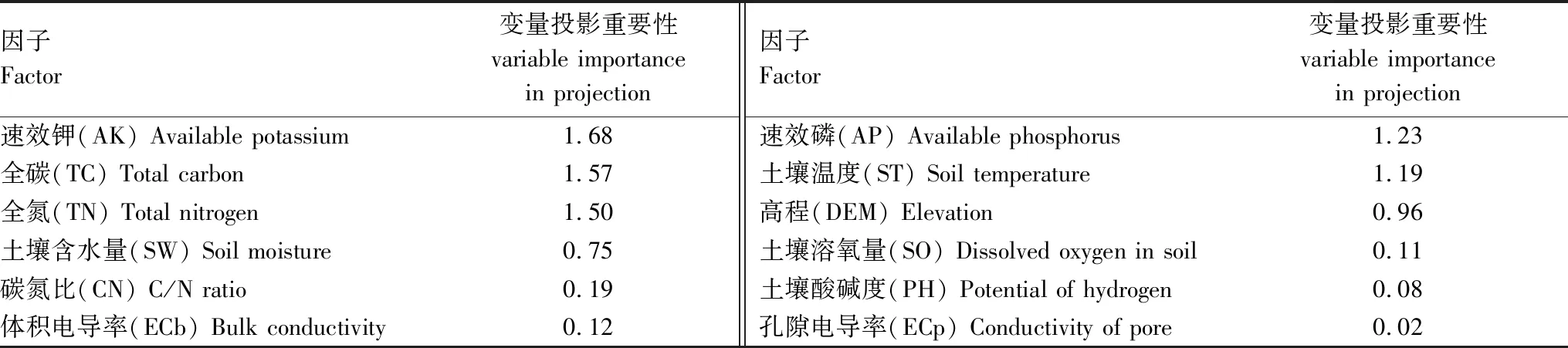
表3 變量投影重要性(蘆葦沼澤)Table 3 Variable importance in projection (Phragmites australis)
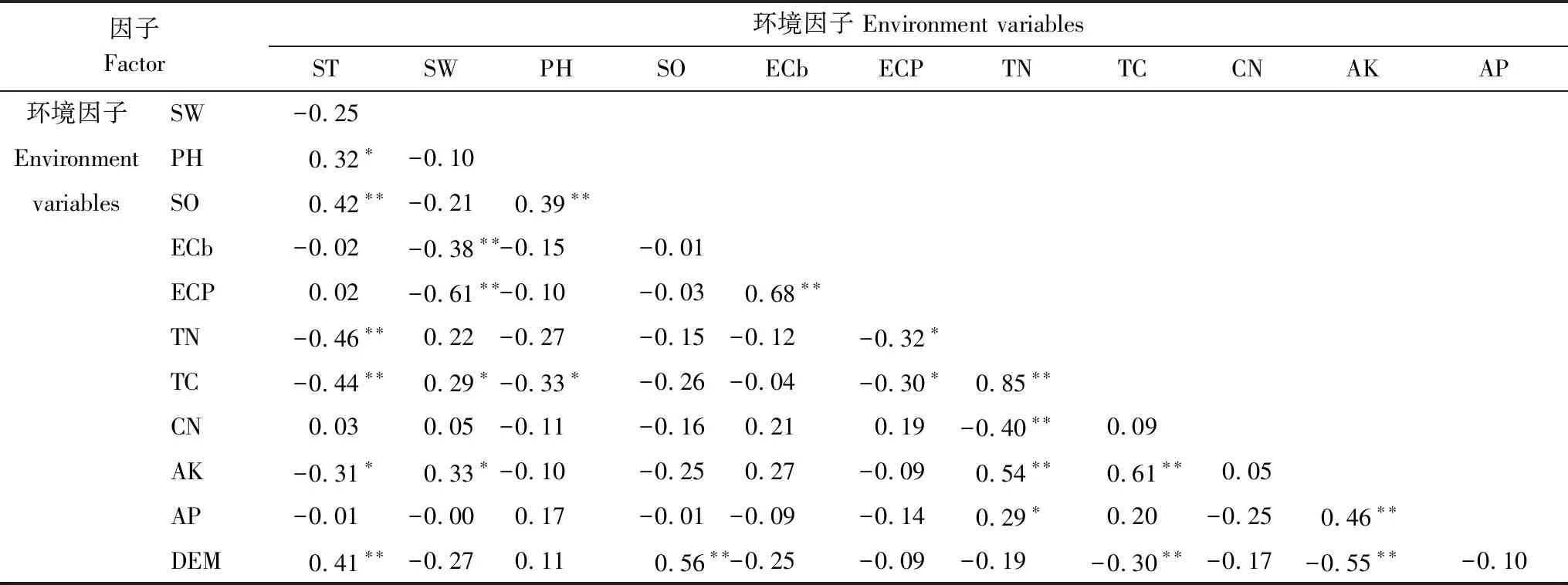
表4 各因子間的相關系數Table 4 The correlations between various factors
3.2.2營養因子對生產力的影響
由圖6可知,N處理組、P處理組和K處理組的平均NPP分別比對照組大7.25%(LRR=0.07)、4.71%(LRR=0.05)和1.22%(LRR=0.01);NP處理組、NK處理組和PK處理組的平均NPP分別比對照組大13.64%(LRR=0.13)、13.43%(LRR=0.13)和6.63%(LRR=0.06);NPK處理組的平均NPP比對照組大15.96%(LRR=0.15)。單個營養限制隨著其他單個營養限制的增加而增加,營養限制隨著第三個營養限制的增加而增加。這說明N、P和K對蘆葦NPP的限制形式是協同限制。濱海鹽沼濕地相對于其他生態系統而言,水鹽的重要性更強且兩者具有較強的相關性(表4),所以在研究NPP營養限制時需要考慮鹽分的作用。如圖7所示,蘆葦NPP的N、P和K限制隨著土壤電導率的增加呈拋物線形式——先增加后較少,但在電導率影響下,各營養限制的敏感性、最大值及其位置存在差異。從拋物線的開口大小可以表明蘆葦NPP在電導率變化條件下對各營養限制的敏感性,開口越小(0.041>0.011>0.008)說明營養限制越敏感。由此可知,K營養限制的敏感性最大,N營養限制最小,P營養限制居中。從拋物線頂點的橫坐標位置說明營養限制最大時的位置,N、P和K營養限制達到最大值時的電導率值分別為4.50、4.36、7.98 ms/cm,可知K限制耐鹽性更強,其次是N和P限制。從拋物線頂點的縱坐標位置說明營養限制的程度,N、P和K營養限制的最大值分別為0.15、0.03、0.34,可知營養限制程度的大小排序依次為K、N和P。
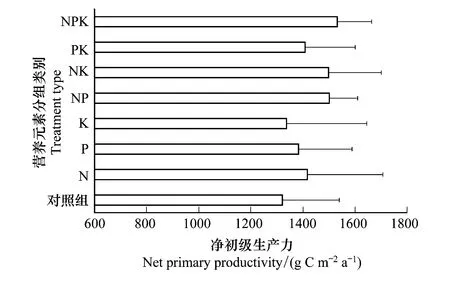
圖6 各分組平均凈初級生產力 Fig.6 The mean net primary productivity with total nitrogen (N), available phosphorus (P), and available Potassium (K) additions
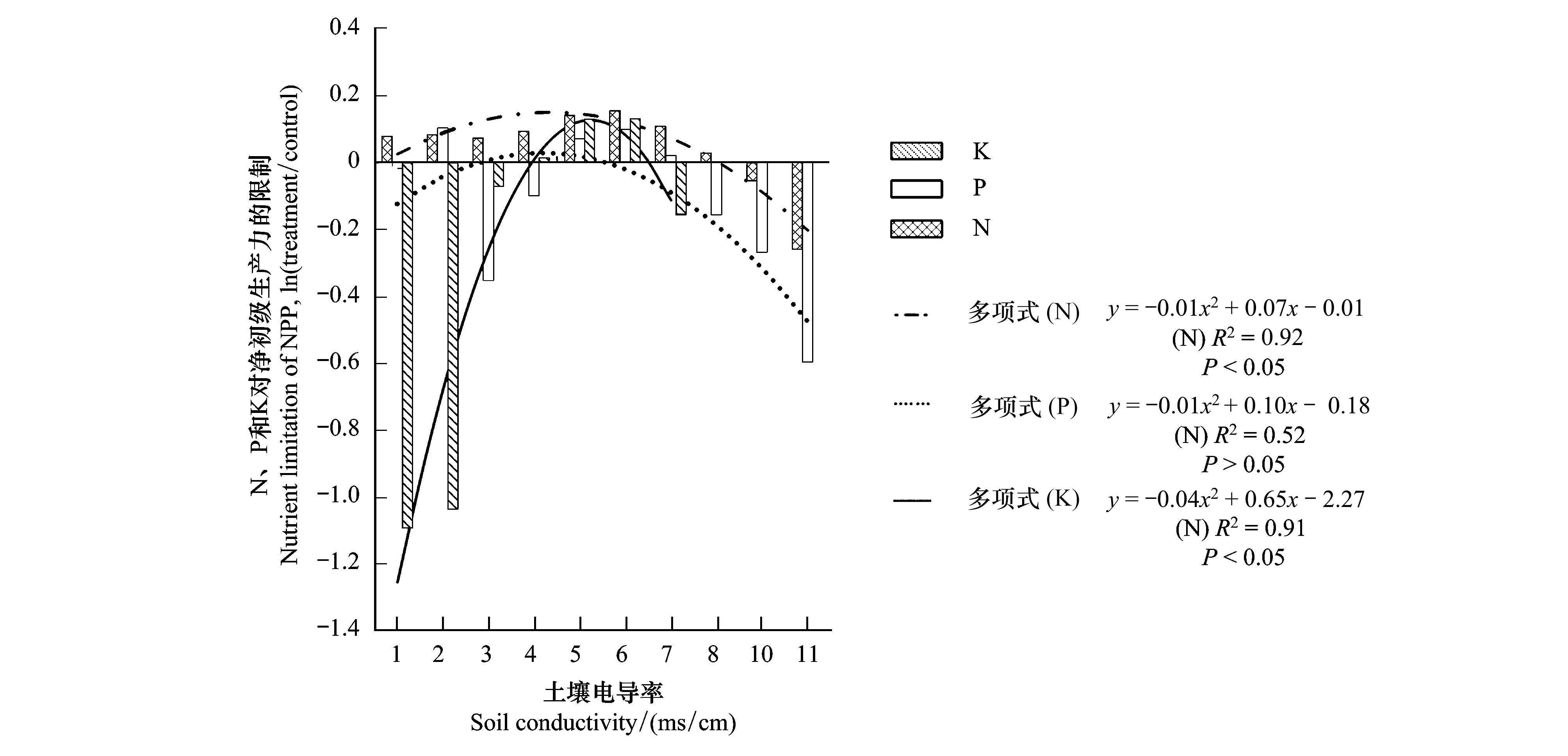
圖7 電導率對N、P和K限制的影響Fig.7 The effect of conductivity on N, P and K limitation
3.2.3土壤水鹽對生產力的影響
土壤水鹽對蘆葦生長的影響也基本符合拋物線模式(圖8),蘆葦NPP隨著電導率或土壤含水量的增加呈先增加后減少的模式。從圖8可以看出,研究區蘆葦NPP對土壤含水量的敏感性小于土壤電導率(9.78>0.26),且土壤電導率對蘆葦NPP的影響程度也大于土壤含水量(最大值1508.52>1493.18 gCm-2a-1);最適的土壤含水量和土壤電導率分別為49.46%和3.96 ms/cm。濱海鹽沼濕地土壤水鹽呈極顯著負相關,相關系數達到-0.38(表4)。不同土壤含水量條件下,土壤電導率對蘆葦NPP的影響也呈近似拋物線模式(圖9)。土壤含水量越高,蘆葦生長對土壤電導率的敏感性越低:土壤含水量34.20%、38.25%和43.40%對應的拋物線二次方系數的絕對值分別為16.35、11.90和6.70,而二次方系數絕對值越大拋物線開口越小,即是相同電導率變化情況下NPP的變化越大。土壤含水量34.20%、38.25%和43.40%對應的最適土壤電導率大小分別為4.28、5.05、2.14 ms/cm,可知并不是土壤含水量越高,最適電導率越大。3種土壤含水量對應最適電導率條件下蘆葦NPP的最大值分別為1523.69、1494.53、1480.79 g C m-2a-1。
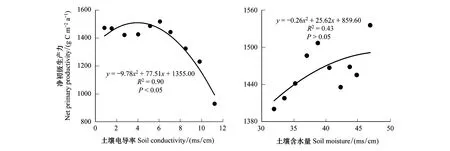
圖8 電導率和土壤含水量與蘆葦凈初級生產力的關系Fig.8 The correlation between conductivity, soil moisture and net primary productivity of Phragmites australis

圖9 不同土壤含水量情況下,電導率對蘆葦沼澤凈初級生產力的關系Fig.9 The correlation between conductivity and net primary productivity of Phragmites australis under different soil moisture
4 討論與結論
4.1 討論
4.1.1鹽沼植被的營養限制
研究表明N、P、K的單獨添加能增加蘆葦NPP(圖6),且NP、NK、PK和NPK對NPP都存在交互效應。單個營養限制隨著其他單個營養限制的增加而增加,營養限制隨著第三個營養限制的增加而增加,表明該這3種營養元素對區域蘆葦NPP具有協同限制效應。Fay等對橫跨五大洲的42個草地監測站點進行養分添加實驗,也表明有31個站點存在營養限制,29個站點存在N、P、K配對組合的協同限制[18]。N、P、K對NPP的平均影響分別是7.25%、4.71%和1.22%,說明N和P是蘆葦NPP的主要限制因子,K是次要限制因子。學者很早就開始了鹽沼植被營養限制的研究[34-35],許多研究得出了鹽沼植被生產力受N和P及NP協同限制的影響[35-38]。本研究也佐證了這一點:對蘆葦沼澤而言,除N和P外,K也是一種限制元素。K限制也被發現存在于泥炭地[39]、沼澤[40]、草地[18]和熱帶雨林[41]。不管怎樣,K限制在一定程度上被忽視了,應在以后的研究中獲得更多的關注。
前人研究也表明蘆葦對K的需求量最大,其次為N和P,土壤N、P、K比例約為13.8∶1∶8.3時,有利于蘆葦生長發育;蘆葦NPP較高時,N、P、K被蘆葦植株吸收的比例約為4.6∶1∶5.3[42]。本研究野外采樣點N、P、K的平均比例約為11.1∶1∶51.7,蘆葦NPP受到K過高的限制。這也表明蘆葦的高產不但對營養元素含量有要求,也會受到營養配比的限制[43]。另外,土壤水鹽對植物的營養限制也起一定的作用。本研究也佐證了這一結論,同時也進一步得出了N、P、K的限制隨土壤電導率的增加可能呈拋物線形式(圖7)。因此,當研究鹽沼植被的營養限制時,需要考慮土壤水鹽影響下的多要素限制。
4.1.2土壤水鹽對鹽沼植被NPP的影響
土壤水鹽是濱海鹽沼濕地形成的重要因子,也是影響鹽沼植被分布格局和生長的重要環境因素。本研究表明蘆葦NPP隨電導率的增加呈先增加后減少的趨勢,基本符合拋物線模式(圖8)。這與邱天等進行的蘆葦水培實驗所得研究結論一致[44]。肖燕等的研究也表明隨著鹽度的升高蘆葦植株高度和分蘗數下降,高鹽抑制了蘆葦的生長,而淡水和中鹽度條件下蘆葦生長較好[45]。Rabie等的研究也表明當土壤鹽度小于等于0.3%時,植被的干物質量隨著土壤鹽度的增加而增加;當土壤鹽度介于0.3%與0.9%時,植被的干物質量隨著土壤鹽度的增加而減少,即干物質量隨土壤鹽度增加呈先增加后減少趨勢[46]。這可能是由于植被都有各自的耐鹽閾值[47],適當鹽分含量對植被的生長具有促進作用,這也從側面佐證了本文的研究結論。
土壤含水量對蘆葦NPP的影響也呈近似拋物線模式(圖8)。這可能的原因是:土壤含水量低一方面會產生氣孔限制或降低蘆葦光系統II(PSII)活性,進而降低蘆葦葉片光合速率,抑制蘆葦生長[48];另一方面會使蘆葦新葉減少,落葉增加、葉片平均大小變小,CO2同化作用和氣孔導度降低,從而導致葉面積和葉生物量減少,最終影響蘆葦NPP[49];隨著土壤含水量的增加,蘆葦的凈光合速率、蒸騰速率、葉片瞬時光能利用效率逐漸增加,最終使蘆葦NPP增加[50]。同時,土壤水分也是影響植被營養元素吸收的重要因子[46]。這說明,在研究鹽沼植被生長的限制因子時,也需考慮土壤水鹽環境的影響。
綜上所述,研究區蘆葦NPP受土壤水鹽和養分的共同影響,不單單是傳統意義上理解的土壤水分含量越高、鹽分越低、養分越充足,蘆葦生長就越好,而可能需要影響蘆葦生長的各個土壤因子達到一種組合最優,才能更好的促進蘆葦生長。
4.2 主要結論
本文以遙感數據、氣象數據和野外采樣數據為基礎,通過模型模擬和統計分析,探究濱海鹽沼濕地土壤因子對植被NPP的影響,主要得到以下主要結論:
(1)研究區NPP空間分異明顯,變異系數為43.71%,植被類型分布整體上決定了區域NPP的空間分異,同時,同種植被類型內部也具有較為明顯的區域分異特征。
(2)土壤理化因子和地形因子對蘆葦NPP影響的敏感性從大到小排序為:AK>TC>TN>AP>ST>DEM>SW>CN>ECb>SO>PH>ECp。
(3)N、P和K對蘆葦NPP的限制形式是協同限制,且該限制隨土壤電導率的增加呈拋物線形式——先增加后較少;在電導率影響下,各限制的敏感性存在差異:K營養限制的敏感性最大,N營養限制最小,P營養限制居中;營養限制程度從大到小排序依次為K、N和P。
(4)土壤水鹽對蘆葦NPP的影響也基本符合拋物線模式:蘆葦NPP隨電導率或土壤含水量的增加呈先增加后減少的趨勢。同時,蘆葦NPP對土壤電導率的敏感性和土壤電導率對蘆葦NPP的影響程度都大于土壤含水量,且土壤含水量越高,蘆葦NPP對土壤電導率的敏感性越低。
(5)只有各土壤因子達到組合最優時,蘆葦NPP才最大。
致謝:感謝國家科技基礎條件平臺-國家地球系統科學數據中心(http://www.geodata.cn)和中國氣象數據網(http://data.cma.cn)提供數據支撐。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
今日農業(2021年11期)2021-08-13 08:53:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
中國塑料(2016年3期)2016-06-15 20:30:00
