浙江天童木本植物水力結構與樹高的關聯性
2020-11-13 09:28:10趙琦琳田文斌史青茹由文輝閻恩榮
生態學報 2020年19期
關鍵詞:植物
趙琦琳,田文斌,鄭 忠,史青茹,3,由文輝,閻恩榮,*
1 云南省生態環境監測中心, 昆明 650034 2 華東師范大學生態與環境科學學院/浙江普陀山海島生態系統研究站, 上海 200241 3 上海福山外國語小學, 上海 200120
垂直分層是森林群落的基本結構特征之一,是植物對垂直空間微環境資源的生態位分化結果。不同垂直層次植物為充分利用光、水資源,在水分生理和解剖結構方面會形成一系列的適應特征[1]。一般而言,群落不同垂直層次植物的植株高度、樹冠疏密度、枝葉關系等構型性狀[2],以及光合速率和蒸騰速率等生理性狀完全迥異[3],反映了植物對光水資源利用方面的功能差異。研究植物的光和水分利用功能在群落不同高度層的屬性,有助于理解植物群落垂直層次結構的成因。
除光照影響外,水分運輸是決定植物生長高度的最主要限制性因素[4]。植物體內的水分運輸途徑包括根系吸收、邊材木質導管輸送、葉片氣孔蒸騰等環節[5]。其中,植物體內水分長距離輸送的動力來源于葉片蒸騰拉力和根壓的共同作用[6]。根據蒸騰拉力-內聚力-張力理論,植物在向樹冠輸送水分過程中,其葉片蒸騰拉力和水分子內聚力顯著高于水柱張力,使得水分不間斷地向冠層運輸[7]。
水力限制是解釋植物水分長距離運輸的另一個重要科學假說,其認為:隨植物高度上升,水分運輸距離增長,運輸阻力和水分重力也會增大。在此情況下,植物必須在輸水效率和安全性方面發生權衡,一方面通過增大木質部導管而提高水分運輸效率,另一方面,為了克服高處植物體內由重力引起的水分較大張力,而引起管道氣穴化,高大植物從基部向上運輸管道逐漸變細,提高水分運輸安全性[4, 8]。因此,植物在生長高度、導管直徑大小和水分傳輸速率間的權衡作用,使得植物高度不可能無限增大[9-12]。從以上推理可以看出,植物的水分傳導與調配方式是影響植物高度和群落層片配置的主要因素。例如:大量的研究通過植物潛在高度與比葉面積、導管數量和干材密度等性狀間的異速關系揭示其內在機制[2, 10, 13-15],但是,關于群落內不同高度植物水分動力特征及其影響因素的理解仍然不夠深入。
本研究聚焦森林群落內部生長于不同垂直層次的木本植物,主要探討木質部水力導度和導管大小等水力結構特征在不同高度植物間的差異。根據以上論述,我們提出以下科學預測:群落不同垂直層次植物的木質解剖和相關的水分動力特征具有顯著差異,高大植物比低矮植物具有更大的水力導度(比導率)。與之匹配,高大植物導管性狀組合策略為疏松排列的大導管,而低矮植物為緊密排列的小導管。為驗證以上預測,本研究以浙江天童木荷群落中生長于不同垂直層次的25個木本植物個體(喬木11株、灌木14株)為研究對象,通過對每株植物木質導管解剖性狀、枝條物理性狀和水分比導率等性狀特征的測量,以期揭示木質部比導率在不同高度植物間是否存在差異,以及植物高度變化引起的導管大小特征是否可以解釋其水力結構特征變化。
1 材料與方法
1.1 研究樣地和樹種選擇
研究地位于浙江寧波天童國家森林公園(29°52′N, 121°39′E),該地屬浙東低山丘陵,土質為紅黃壤,成土母質為中生代的沉積巖、部分酸性火成巖以及石英砂巖和花崗巖的殘積風化物。土壤厚薄不一,一般在1 m以下。氣候屬于亞熱帶季風氣候,雨量充沛,年平均氣溫16.2℃,年平均降雨量1389.7 mm,年無霜期230—240 d。主要植被類型為常綠闊葉林,其中木荷(Schimasuperba)林分布面積最多[16]。
本研究對象為位于天童國家森林系統觀測研究站西側100 m處的長期固定樣地外圍的木荷群落。樣地海拔167 m,東北坡向,坡度25°。群落垂直分層明顯,喬木層高度16 m左右,蓋度80%;優勢種為木荷,另有少量石櫟(Lithocarpusglaber)。灌木層最大高度5 m,蓋度60%;主要種類有馬銀花(Rhododendronovatum)和連蕊茶(Camelliafraterna)等。草本層主要為里白(Hicriopterisglauca)等。
為探索群落內不同垂直層次共生木本植物的水力結構特征,本研究在該樣地選擇了高度不同的25個木本植物個體,其隸屬于9個物種,高度大于10 m以上的個體11株,小于10 m的個體14株(多數集中在5.5m以下),詳細情況見表1。

表1 浙江天童木荷群落研究樹種特征及其邊材面積和胸徑的回歸模型Table 1 Characteristics of selected species and regression equation between sapwood area (A) and diameter at breast height (DBH) for each species in Schima superba community in Tiantong, Zhejiang Province
1.2 植株高度、枝條導水率和比導率的測定
野外和室內測量于2014年7—9月開展。植株高度利用樹木高度測量儀(Vertex—IV, Hagl?f, Dalarna, 瑞典)完成。枝條比導率測量選擇晴朗天氣進行。測量期內,于7:00采集待測植株樹冠頂部生長良好的枝條。為消除樹冠不同受光方向枝條間的差異,采集枝條時,從3個不同方位剪取具有多級分枝的枝條,作為重復[9]。在后續的水力和解剖結構參數測量時,從多級分枝中專門挑選二年生枝作為對象,這是由于二年生枝條木質結構發育較為完整,其強度也滿足后續各測試環節操作需要。為盡量減少不同部位枝條大小差異的影響,所選擇的枝條直徑大小保持一致[12]。在野外,待枝條剪去后,立即將其浸沒于水桶,并在水下切口,以防止氣泡進入枝條導管而發生空穴化。同時,用黑色塑料袋罩住待測枝條,以減少光照輻射,避免蒸騰。完成以上程序后,立即將樣品帶回實驗室。
在室內,將選取的枝條在水下分割為10—20 cm的枝段,首先采用HPFM—Gen3植物導水率高壓測量儀測定導水率。測量前,先用去氣蒸餾水注入高壓測量儀,然后用大排水管和毛細管,在450 kPa恒定壓力下排氣12 h以上。樣品測量開始時,將壓力調零,連接枝段末端與壓力耦合器,以壓強10 kPa為起點,按5—10 kPa/s的速率,逐漸加壓到500 kPa左右,從而得到壓強和流速變化關系的回歸曲線,枝條導水率用該曲線的斜率來表征。
完成以上工作后,用刀片刮去樹皮,測量枝段半徑。之后,對其進行有色溶液染色,在顯微鏡下測量邊材直徑,進而推算邊材面積(有效輸導面積)。最后,根據導水率除以邊材橫截面積得到比導率[17]。
1.3 木質導管大小和枝條密度測定
帶回實驗室的預留枝條用于導管直徑測量。首先,將枝條切割為2—4 cm的片段,并立刻保存在FAA固定液中。導管測量前,用滑走切片機將保存的小枝段橫向切割為厚度20—30 μm的切片,選取完好的移至表面皿中。然后,用蒸餾水充分清洗,再用乙醇逐級多次脫水,根據苯胺番紅—苯胺固綠雙重染色法充分染色。最后,再用95%酒精洗去切片浮色,進一步進行乙醇脫水,完成二甲苯透明后,將切片小心固定在載玻片上,利用中性樹膠封存載玻片。
完成以上步驟后,將載玻片安裝在數字照相顯微鏡(Olympus DP73, 日本)中測量導管的分布和尺寸。測量時,將顯微鏡放大20倍,首先選擇一個視野,根據木質射線邊界和導管管腔影像確定導管大小和數量,然后依次增加觀察視野數,直至記錄到40個導管為止。以上過程中,每個視野都進行拍照保存,然后使用自動分析軟件(Image—Pro Plus 7.0C, America)統計平均導管直徑、平均管腔面積與給定視野面積中的導管數量。最后,根據平均導管直徑、給定視野面積中的導管數量和枝條橫截面積的乘積,得到枝內腔面積,該參數一定程度上也能反映了枝條的導水效率[18]。
另外,選取帶回的大枝測量材質密度。每個植株也測量3個枝條作為重復。測量時,先刮除樹皮,稱量鮮重后,采用排水法測量體積。之后,將其置于70℃烘箱中烘干至恒重(一般48 h),稱量干重。枝條密度為單位體積的干物質份數[14]。
1.4 數據處理
由于研究個體的高度界限明顯,為了探索是否上層木和下層木類別的水力結構特征具有顯著差異,因此首先將25個個體以10 m為界歸為兩組,各層所有個體的測量參數取平均,按照配對樣本T檢驗(Paired-Ttest),分析了枝條比導率、導管密度、直徑和內腔面積,以及枝內腔面積、枝密度、枝直徑和邊材面積在上層和下層木間差異顯著性。其次,利用一元線性回歸,分析各水力結構性狀和物理性狀是否隨樹高而顯著變化;同時,分析比導率是否與其他解剖學性狀和物理性狀間具有顯著關聯性。
2 結果
2.1 植物水力結構和物理性狀與樹木大小的關系
上層木的比導率顯著高于下層木(表2),表明前者的水分傳導效率顯著大于后者。干材密度和木質導管密度在兩者間的差異不顯著(P>0.05)(表2)。下層木的導管直徑和內腔面積、枝直徑和內腔面積以及邊材面積均顯著小于上層木(P<0.01)(表2)。

表2 上層和下層木植物枝條比導率、木質導管和邊材性狀的差異Table 2 Differences in specific conductivity of twigs and xylem vessel and sapwood traits between overstory and understory plants
線性回歸分析顯示,導管直徑和內腔面積、枝條直徑和內腔面積和邊材面積,以及枝條最大導水率均與樹高間顯著正相關(圖1)。
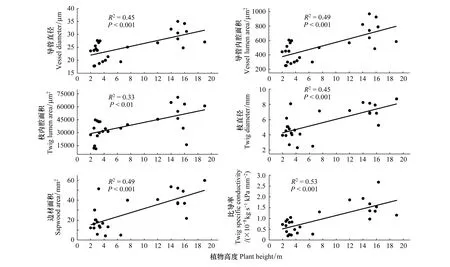
圖1 枝條物理和水力性狀與植物高度的回歸關系Fig.1 Regression of plant height with each of physical and hydraulic traits
2.2 植物枝條比導率與木質解剖和物理性狀的關系
比導率與導管直徑和內腔面積、枝直徑和內腔面積、邊材面積以及枝條最大導水率均顯著正相關(圖2)。干材密度和導管密度均與比導率無顯著相關性(P>0.05)。
3 討論
3.1 森林群落不同垂直層次植物水力結構特征的分化
以浙江天童木荷群落內的25株生長于不同高度的木本植物為對象,通過分析枝條水分比導率等水力結構參數和木質導管等性狀與樹高的關系,本研究結果表明,上層木的枝條水分比導率顯著高于下層木,導管直徑和內腔面積、枝條直徑和內腔面積,以及枝條邊材面積和最大導水率均隨樹高增大而增加。因此,該結果與本研究的科學假說一致,即:對生長于木荷群落不同垂直層次的木本植物,其木質部導水性能及其相關聯的木質導管性狀也具有顯著的差異。本研究中的木荷群落處于常綠闊葉林演替中后期[16],群落結構趨于穩定,植物枝條水力結構特征表現出的明顯垂直梯度變化規律,反映了不同垂直層次植物的多樣化水力結構策略[19]及其水分生態位分化過程[20]。
喬木層個體多數為木荷,作為演替亞頂極種,占據群落上木層,樹冠開闊緊湊,枝葉濃密,垂向枝條比例遠大于橫向枝比例,具有極大的獲取空間和光水資源的優勢[1]。與之相比,處于林下的物種多為常綠闊葉林耐蔭灌木,比如馬銀花和連蕊茶等,其潛在最大生長高度在5 m左右,垂向水分運輸成本較低[21];其次,這些物種一般冠形松散,橫向枝比例較大,其在捕獲林下光斑方面具有優勢,但不利于垂向的水分運輸[2, 22]。另外,相較于林下灌木,處于森林上木層的個體在獲取光資源方面占據優勢的同時,其必然也要付出較高的生理代謝和水分利用效率成本[23-24],這樣的需求必然需要依靠更大的木質部水分比導率[25]和更大的導管內腔面積來維持[18, 26]。
更為重要的是,在水分運輸距離方面,生長于群落上木層的植物要比下木層植物付出更大的代價[27-28]。垂向水分運輸動力來自蒸騰拉力和根壓的共同影響[6]。通常,對于高大喬木,根壓的貢獻相對較小,蒸騰拉力是驅動水分長距離運輸的主要動力。研究表明,樹高與樹冠總的蒸發量成正比[29],蒸騰作用會通過葉表氣孔蒸發消耗大量水分,從而在葉片、枝條、樹干直至根部形成一個極大的水勢梯度[30],從而保障水分有效運輸到高大喬木的樹冠部位[4, 8]。保障水分長距離運輸的其他動力還有水柱與管壁間的附著力以及水分子間的內聚力[31-32]。
3.2 不同高度植物水分比導率與導管解剖特征的關聯性
在本研究中,干材密度在喬灌木間差異不顯著。但是,決定干材密度的導管大小和內腔面積在喬灌木間差異顯著,該現象與加利福尼亞州207個木本植物導管差異的比較結果一致[33],即:與灌木相比,喬木具有大的導管直徑和內腔面積。另外,本研究也發現,導管直徑和內腔面積對不同高度植物枝條水分比導率的差異具有較高的解釋率(圖2)。過去的研究表明,在植物的不同發育階段,導管直徑會隨個體發育而增加,這樣,導管寬度的增大可以抵消植株高度增大帶來的水力阻力[9-10, 27-28]。與灌木相比,喬木擁有大的導管直徑則有利于克服長距離水分運輸的生物物理限制。以上研究綜合表明,群落不同垂直層次個體水分比導率等水力結構的差異可從其木質導管的大小變化得以解釋[3, 25, 33]。
喬灌木導管大小變化的可能原因如下。喬木比灌木的水分運輸距離更長,需要克服的運輸阻力和重力更大,那么就需要更大的導管來提高水流通量來抵消。這個水力學機制可從泊肅葉定律(Poiseuille′s law)得到證明,即:植物液流通量與導管的半徑四次方成正比,而與水分輸導距離為反比。對于任何植株而言,要維持一個固定的液流通量,那么,每增加1個單位的水分運輸距離,就需要導管半徑按四次方的倍數增加才可達到平衡。由于植物液流通量與枝條水分比導率顯著正相關[31],顯然,這就可以理解為什么高大植物傾向于選擇更大的導管直徑。
需要特別提及的是,本研究發現導管直徑和內腔面積與比導率顯著正相關,而導管密度與水分輸導性狀無顯著關系。一般而言,導管密度反映了木質部維管系統的疏密程度,排列緊密的導管不利于增強導水率,但有利于減少空穴化和栓塞化[31,33]。研究表明,雖然導管大小可影響導水率[7],但導管密度對邊材導水率影響較小[32],其具體原因還有待進一步研究。
總之,本研究通過研究一個局域群落內共存于不同垂直高度的植物個體的水力學和導管解剖學特征,揭示了樹高是影響是植物水分利用、運輸策略的重要因素,其中,木質導管大小變化是決定不同高度層植物水力結構的重要因素。本研究結果強調了群落不同層片植物水力結構策略的權衡是維持不同垂直層次物種共存的重要機制。另外,由于各方面的限制,本研究所涉及的群落和物種數較少,未來還需要進一步擴大研究對象的樣本數,力求得到更加普遍性和更深入的結論。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
