豬β-防御素1在畢赤酵母中的串聯表達
2020-09-29 08:39:34范闊海孫耀貴李宏全
中國獸醫雜志 2020年4期
王 鑫,范闊海,尹 偉,孫 娜,孫耀貴,李宏全
(山西農業大學動物科技學院,山西 太谷 030801)
自青霉素問世以來,人們對病原菌造成的感染性疾病不再束手無策,但是由于細菌對抗菌藥物的廣泛抑制,發現和開發新型有效的抗菌劑就成為了全世界備受關注的焦點。天然產生的抗菌肽(Antimicrobial peptides,AMPs)廣泛分布于整個動植物界,具有分子量小、熱穩定性好、廣譜抗菌和作用靶點為病原體細胞膜[1-2]等特點,顯示出高效的殺菌能力,被認為是未來最有希望的代替者[3]。防御素(Defensin)是一類在動植物體內廣泛存在[4]、分子量大約3~5 kDa,具有抗細菌[5]、真菌[6]、病毒[7]等微生物生物學活性的內源性陽離子抗菌肽,主要來源于上皮組織,是非特異性天然免疫系統的重要組分之一,在腸道穩態和抵抗病原體方面起著至關重要的作用[8]。
豬源防御素家族的結構比較保守[9],富含半胱氨酸,多聚陽離子型。豬β-防御素可通過直接殺菌作用抑制細菌感染,同時抑制促炎細胞因子的釋放減輕炎癥反應[10]。高表達β-防御素的豬可抵抗豬病原性胸膜肺炎放線桿菌的感染[11]。豬β-防御素1(Porcine beta-defensin1,pBD1)是豬體內最早發現的防御素,在豬的全身組織及器官中廣泛表達,不同品種及不同的生長階段的豬其表達情況也不盡相同。第1個pBD1基因由Zhang G L等[12]克隆出來,編碼64個氨基酸,成熟肽有38個氨基酸。pBD1在大腸桿菌表達系統和昆蟲桿狀病毒表達系統中的表達已有報道。姜麗華等[13]首次在畢赤酵母表達系統中實現了pBD1的分泌表達,表達產物對金黃色葡萄球菌有較好的抑菌活性,但是表達量較低不適合大規模生產。本試驗為了高效表達pBD1蛋白,設計合成pBD1三倍串聯基因(3pBD1),構建重組表達載體pHLI-S1-3pBD1,建立巴斯德畢赤酵母(Pichiapastoris)對3pBD1的真核表達系統,為pBD1的進一步大量生產并開發成新型抗菌藥提供理論基礎和技術路徑。
1 材料與方法
1.1 菌株及載體 畢赤酵母宿主菌GS115、大腸桿菌TOP10、酵母表達載體pHIL-S1均為本實驗室保存。
1.2 主要試劑 基因組DNA提取試劑盒,購自北京索萊寶科技有限公司;TRIzol Reagent總RNA提取試劑盒,購自美國Invitrogen;E.Z.N.A.TMGel Extraction Kit、E.Z.N.A.?Plasmid Mini Kit I、Prime Script RT-PCR Kit、EcoRⅠ、BamHⅠ限制性內切酶均,均購自寶生物工程(大連)有限公司。
1.3 方法
1.3.1pBD1三倍串聯基因的設計 根據GenBank中pBD1基因(NM_213838.1)的成熟肽序列和巴斯德畢赤酵母密碼子的偏好性設計3pBD1,設計的3pBD1基因全長625 bp,包括202個氨基酸編碼區、終止密碼子、2個腸激酶識別位點、EcoRⅠ酶切位點和BamHⅠ酶切位點,將設計好的序列送生工生物工程(上海)股份有限公司合成。3pBD1基因所編碼的氨基酸共有202個,分子量約為22 kDa。
1.3.2 重組表達載體pHIL-S1-3pBD1的構建及鑒定 將目的基因3pBD1和表達載體pHIL-S1用BamH Ⅰ/EcoRⅠ進行雙酶切反應,雙酶切反應體系為:EcoRⅠ 1 μL,BamHⅠ 1 μL,K-Buffer 2 μL,載體6 μL,ddH2O補齊至20 μL。37 ℃酶切12 h,2%瓊脂糖凝膠電泳檢測,割膠回收。將上述回收的pHIL-S1和3pBD1進行連接,連接反應體系為:pHIL-S1載體1 μL,3pBD1基因7 μL,T4 DNA連接酶1 μL,10×T4 DNA Buffer 1 μL。利用熱激法將連接產物轉化至感受態大腸桿菌TOP10,涂布于含有Amp(100 mg/mL)的LB平板37 ℃培養12 h。
挑取單菌落接種于5 mL含有Amp(100 mg/mL)的液體LB培養基中,37 ℃,180 r/min培養12 h。提取載體使用BamHⅠ/EcoRⅠ進行雙酶切鑒定并利用通用引物3′-AOX1Primer、5′-AOX1Primer進行PCR鑒定,篩選出陽性重組表達載體。
1.3.3 重組表達載體pHIL-S1-3pBD1的酵母電擊轉化 線性化重組表達載體pHIL-S1-3pBD1,反應體系為:SalⅠ1 μL,10×L Buffer 2 μL,重組載體4 μg,加ddH2O至20 μL。37 ℃酶切12 h,4 ℃終止酶切,2%瓊脂糖凝膠電泳檢測,以線性化的空pHIL-S1作對照。
制備GS115酵母感受態細胞,取畢赤酵母GS115菌種劃線接種于YPD平板上,30 ℃過夜培養。挑取單菌落接種于5 mL YPD培養液中,30 ℃,250 r/min培養12 h。加0.5 mL上述畢赤酵母菌液到50 mL新鮮培養基中,30 ℃,250 r/min培養至OD600值達0.8~1。以下步驟在冰上操作:取50 mL菌液于預冷的50 mL離心管中,1 500 g,4 ℃,離心5 min,棄去上清。加50 mL冰水于沉淀菌中,重新懸浮,1 500 g,4 ℃,離心5 min,棄去上清。加25 mL冰水于沉淀菌中,重新懸浮,1 500 g,4 ℃,離心5 min,棄去上清。加2 mL預冷1 mol/L山梨醇于沉淀中,重新懸浮,1 500 g,4 ℃,離心5 min,棄去上清,加0.1 mL冰水于沉淀菌中,重新懸浮,立即使用。
電轉化重組表達載體pHIL-S1-3pBD1,取20 μL線性化重組載體加入80 μL酵母感受態細胞,立即放入0.2 cm冰電轉杯中。冰浴5 min。依照電轉儀說明書,以電容25 μF、電壓1.5 kV、電阻200 Ω 進行電擊,電擊結束后立即加1 mL 1 mol/L的冰浴山梨醇于電轉杯中,并將其轉移到無菌的1.5 mL離心管中。取200 μL涂布于MD板。30 ℃靜置60 min,待液體全部吸收后,倒置培養3 d,直到轉化子出現。同時取空載體pHIL-S1作相同處理,用于構建背景對照菌株。
按照北京索萊寶科技有限公司基因組提取試劑盒說明書提取重組酵母基因組。以基因組為模板,用pHIL-S1載體測序引物5′AOX1 Primer和3′AOX1 Primer進行PCR反應。1%瓊脂糖凝膠電泳檢測,送往TaKaRa測序。
對重組轉化子進行Mut表型鑒定。把涂布到MD平板上生長出來的酵母單菌落挑出,采用針點接種的方式將同一陽性菌落分別接種于MM平板和MD平板,盡量控制接種菌量一致,且接種時不要涂布,用針尖點種,保證接種面積相同,30 ℃倒置培養3 d,根據菌落的大小與生長速度判斷轉化子的表型。在MM平板與MD平板上均能正常生長且菌落大小相當的轉化子為Mut+表型;在MD平板上生長正常,在MM平板上幾乎不能生長或生長很緩慢,則為Muts表型。
1.3.4 重組畢赤酵母的誘導表達 將篩選出的Mut+型陽性重組表達菌株接種于25 mL BMGY培養基中,30 ℃,250 r/min振搖至OD600值達到2~6時,室溫3 000 r/min離心5 min收集菌體;用5 mL的BMMY重懸菌體傳于200 mL BMMY培養基中,30 ℃,250 r/min振搖。于培養24、48、72和96 h時補加甲醇至終濃度0.5%。同時以空載體轉化的宿主菌作對照。誘導培養96 h后4 000 r/min 離心10 min收集培養上清液,真空凍干后進行SDS-PAGE檢測。
2 結果
2.1 目的基因3pBD1的獲取 利用EcoRⅠ和BamHⅠ對pUC57-3pBD1進行雙酶切,凝膠電泳檢測625 bp處有清晰條帶(圖1),與設計的目的基因3pBD1大小一致,割膠回收純化獲得3pBD1基因。

圖1 EcoRⅠ和BamHⅠ 酶切pUC57-3pBD1瓊脂糖凝膠電泳分析Fig.1 Agarose gel electrophoresis analysis of pUC57-3pBD1 restriction digestion by EcoR Ⅰ and BamH ⅠM:DL-2 000分子量標記;1~3:酶切產物M:DL-2 000 Marker;1~3:Restriction digested products
2.2 重組表達載體pHLI-S1-3pBD1的鑒定 瓊脂糖凝膠電泳結果顯示(圖2):重組載體pHIL-S1-3pBD1經EcoRⅠ和BamHⅠ雙酶切后,在8.3 kb和625 bp處各有1條清晰的電泳條帶,表達載體pHIL-S1無625 bp的條帶,證明3pBD1基因已經連接至pHIL-S1。
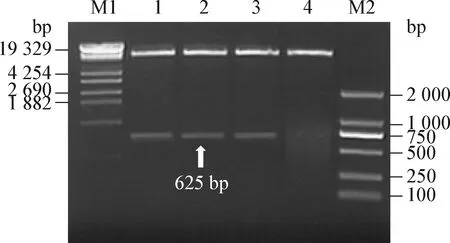
圖2 pHIL-S1-3pBD1酶切瓊脂糖凝膠分析Fig.2 Restrictive endonuclease mapping of pHIL-S1-3pBD1M1:分子量標記;M2:DL-2 000 Marker;1~3:重組載體的EcoRⅠ/BamHⅠ 雙酶切;4:提取重組載體/未酶切M1:Molecular weight marker;M2:DL-2 000 Marker;1~3:Agarose gel electrophoresis analysis of recombinant vector digested with EcoR Ⅰ/BamHⅠ;4:Extract of recombinant vector/undigest
2.3 重組酵母的鑒定 瓊脂糖凝膠電泳結果顯示(圖3):重組酵母菌的基因組經PCR擴增后,在850 bp處有1條清晰的電泳條帶,空載體和空酵母菌無850 bp的條帶,證明重組載體pHIL-S1-3pBD1構建正確并成功導入畢赤酵母中。并且重組酵母PCR產物測序結果與人工設計的3pBD1基因序列同源性為99%,進一步充分說明重組載體pHIL-S1-3pBD1構建正確并成功導入畢赤酵母中。
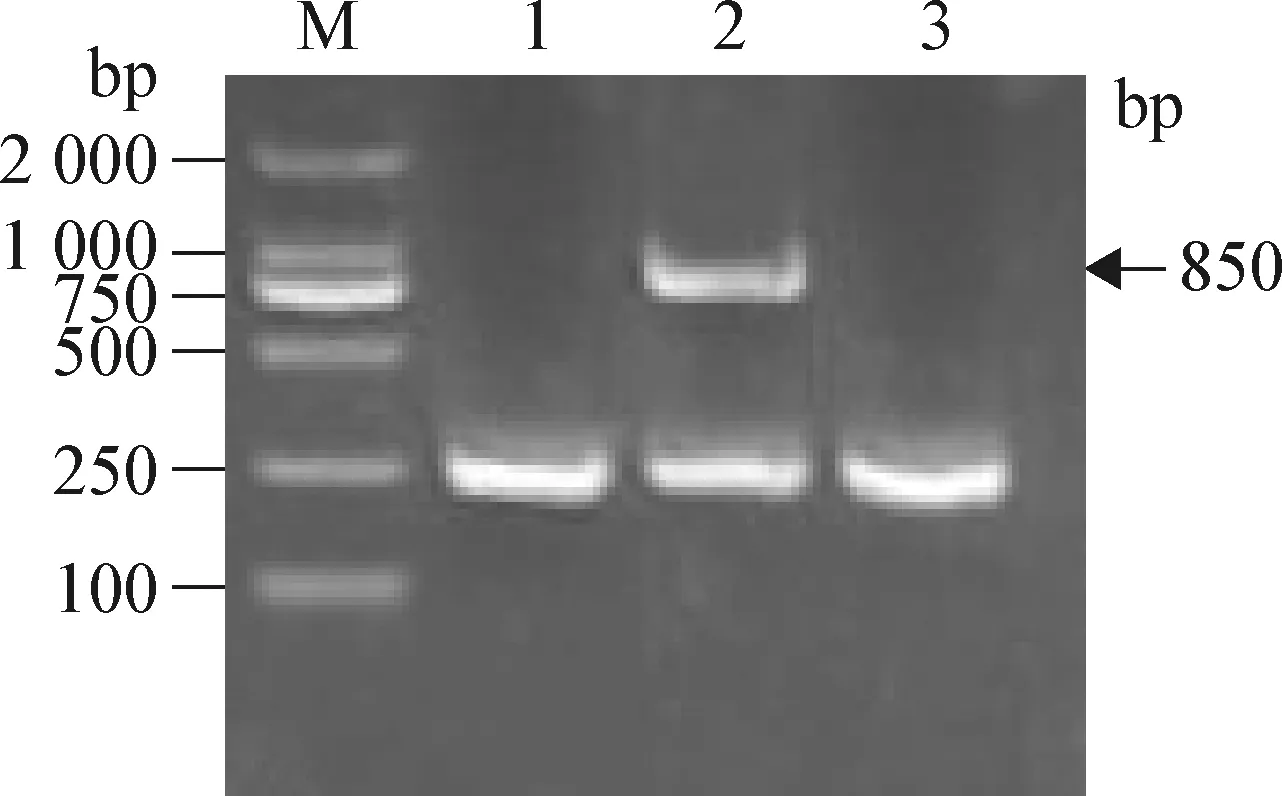
圖3 重組酵母PCR鑒定瓊脂糖凝膠電泳分析Fig.3 Agarose gel electrophoresis analysis of GS1115/pHLI-S1-3pBD1M:DL-2 000分子量標記;1:空穿梭載體PCR;2:重組酵母PCR產物;3:空酵母菌PCRM:DL-2 000 Marker;1:PRC production of pHLI-S1;2:PRC production of GS1115/pHLI-S1-3pBD1;3:PRC production of GS1115/pHLI-S1
2.4 陽性轉化子的Mut表型鑒定 Mut表型鑒定顯示(圖4):獲得的重組酵母菌株在MM平板與MD平板上均能正常生長且菌落大小相當,為Mut+表型。

圖4 陽性轉化子的表型鑒定Fig.4 Identification of mutphenotype of the positive transformants
2.5 表達產物的SDS-PAGE電泳 SDS-PAGE檢測結果顯示(圖5):重組酵母菌株經甲醇誘導表達后,在22 kDa處有1條清晰的特異條帶,含有空載體pHIL-S1的酵母菌株在22 kDa處無條帶,證明3pBD1基因在畢赤酵母酵母中得到了表達。
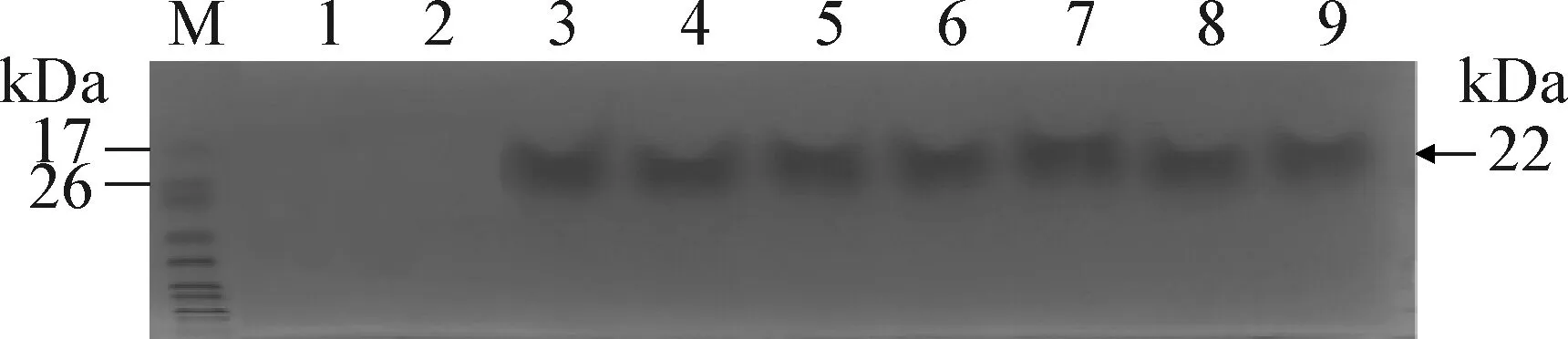
圖5 3pBD1表達上清SDS-PAGE結果Fig.5 SDS-PAGE results of culture supernatant of 3pBD1M:蛋白分子量標記;1~2:GS1115/pHLI-S1;3~9:96 h的表達上清濃縮后產物M:Molecular weight markers for protein;1~2:GS1115/pHLI-S1;3~9:Concentrated sample of 96 hours respectively
3 討論
畢赤酵母表達系統具有獨特的優勢[14-15],作為真核表達系統,可對表達的蛋白進行正確的加工折疊和翻譯后修飾、外源基因通過載體整合到畢赤酵母基因組上,得到菌株比較穩定、外源蛋白既可存在于胞內又可分泌到胞外、生長速度快、表達量高、培養成本低、糖基化程度低、背景蛋白少,畢赤酵母作為外源蛋白表達系統投放市場以來,經歷了許多科研工作者的改進和優化,有極大的應用前景[16]。
防御素是一類對抗外源病原體入侵的肽類物質,是動物自身免疫防御系統的重要組成部分[17]。由于天然抗菌肽在生物體內的含量低,并且體外抑菌效果有限,本試驗將串聯技術與融合表達技術結合起來,根據酵母密碼子偏好性設計了pBD1的三倍串聯體3pBD1基因,適當增加拷貝數,提高目的蛋白的表達效率,且各肽段間加入腸激酶切割位點(DDDDK)以獲得活性蛋白質單體,與分泌型真核表達載體pHLI-S1連接構建重組表達載體pHIL-S1-3pBD1,SalⅠ線性化后電轉至畢赤酵母宿主菌GS115,對重組轉化子進行Mut表型鑒定,篩選得到Mut﹢表型進行甲醇誘導表達,實現了小分子多肽在體外的高效表達。
本試驗選用的受體菌是Cregg建立的GS115。其表型為His-Mut+,其染色體基因組中HIS4(編碼組氨酸脫氫酶的基因)發生了突變,因而不能合成組氨醇脫氫酶,只能在含有組氨酸的培養基中生長。載體pHLI-S1含有與GS115染色體DNA同源的乙醇氧化酶(Alcohol oxidase,AOX1)序列和完整的HIS4基因,通過同源序列的介導,兩者可以發生同源重組,HIS4處的突變被修復,重組菌株可以自行合成組氨醇脫氫酶,故可在組氨酸缺陷的MD培養基上篩選。AOX1是甲醇代謝途徑的第1個限速酶,AOX1基因的啟動子是甲醇誘導型強啟動子,常用于表達外源基因[18]。當SalⅠ線性化的重組載體pHIL-S1-3pBD1轉化宿主菌GS115時,理論上一般在HIS4基因位點進行整合,此時由于AOX1的存在,得到的轉化子原則上都是Mut+,它可以在以甲醇為唯一碳源的MM培養基上正常生長。但由于線性化的重組載體也有可能在AOX1位點發生整合,當在該位點整合時AOX1基因就會因外源基因的插入而失活或被外源基因替換,此時酵母染色體上雖然存在與AOX1具有97%同源性的AOX2基因,但其利用甲醇作為碳源的能力遠遠低于AOX1,這樣轉化子在以甲醇為唯一碳源的MM培養基上生長十分緩慢,即表現為Muts。本試驗中所有的陽性轉化子均為Mut+型。將Mut+型陽性重組表達菌株經BMGY和BMMY培養基培養,甲醇誘導表達96 h后,經SDS-PAGE檢測,在22 kDa處得到1條特異性條帶,而在含空載體的GS115對照處沒有條帶,初步證明3pBD1基因在畢赤酵表達系統中得到了表達,為后期培養表達條件的優化和大規模生產提供了科學依據。
