miR-224靶向抑制ECRG4表達對非小細胞肺癌細胞增殖、遷移和侵襲的影響
2020-09-14 09:00:34趙寅生吳曉燕
國際檢驗醫學雜志 2020年17期
張 輝,趙寅生,吳曉燕
(1.青海省心腦血管病專科醫院胸外科,青海西寧 810012;2.青海省交通醫院病理科,青海西寧 810008)
非小細胞肺癌(NSCLC)占所有肺癌的80%[1]。微小RNA(miRNA)是一類非蛋白編碼RNA,可通過負調節信使RNA的穩定性或抑制其翻譯來實現轉錄后對基因的表達調控[2]。研究表明,miRNA在細胞的生長、凋亡、分化和發育過程中起重要的作用[3]。此外,miRNA的表達譜被證實在肝癌、肺癌等多種腫瘤中具有原癌基因或抑癌基因的功能[2]。據報道,miR-224在乳腺癌、膽管癌和NSCLC中高表達[4-6]。然而,miR-224在NSCLC中的作用和分子機制仍未完全闡明。食管癌相關基因4(ECRG4)位于染色體2q12.2處,最初被認為是食管癌中的抑癌基因[7]。有研究表明,ECRG4在其他多種腫瘤中也是候選的抑癌基因[8]。最近研究表明,自噬基因Beclin-1可通過上調ECRG4通路,抑制A549肺癌細胞增殖,并促進其凋亡[9-10]。然而,ECRG4在NSCLC中的作用機制仍不明確。本研究通過探討miR-224、ECRG4在NSCLC中的表達,以及miR-224靶向ECRG4對NSCLC細胞A549增殖、遷移和侵襲的影響,來進一步揭示NSCLC發生、發展的分子機制,以期為臨床診斷治療提供新的思路。
1 資料與方法
1.1一般資料 收集2016年1月至2018年12月于青海省心腦血管病專科醫院行手術切除治療的NSCLC患者癌組織和癌旁組織標本共68例,其中男39例,女29例;年齡37~75歲,平均(48.5±11.2)歲;高分化28例,中低分化40例;TNM分期Ⅰ~Ⅱ期25例,Ⅲ~Ⅳ例43例。所有患者均經病理診斷證實為NSCLC,且術前未接受任何放療或化療。本研究經青海省心腦血管病專科醫院倫理委員會審核批準,所有研究對象或其家屬均簽署知情同意書。
1.2儀器與試劑 Lipofectmine 2000轉染試劑(批號:11668019)購自美國Invitrogen公司;RNAiso plus(批號:9108)、PrimeScriptTMRT reagent Kit with gDNA Eraser(批號:RR047A)、SYBR?Premix Ex TaqTM(批號:DRR081A)、TakaRa點突變試劑盒(批號:R401)均購自大連TaKaRa公司;放射免疫沉淀法(RIPA)裂解液(批號:C1053)、二喹啉甲酸(BCA)蛋白定量試劑盒(批號:P0009)均購自上海碧云天生物技術有限公司;Dual Luciferase?Reporter Assay System試劑盒(批號:E1960)購自美國Promega公司;anti-miR-NC、anti-miR-224、si-NC和si-ECRG4均購自廣州銳博生物技術有限公司;ECRG4(批號:ab224077)、GAPDH(批號:ab181602)抗體均購自美國abcam公司;5′和3′-雙地高辛標記的MIRURY LNA microRNA Detection Probe試劑盒(批號:10000-89999-15)購自EXIQON公司。健康人肺支氣管上皮細胞系HBE和NSCLC細胞系H460、A549、L78購自中國科學院典型培養物保藏委員會細胞庫,于實驗室的液氮罐中保存備用。
1.3方法
1.3.1細胞培養與轉染 HBE、L78用含10%胎牛血清的DMEM培養基培養,A549用含10%胎牛血清的F-12k培養基培養,H460用含10%胎牛血清的RPMI 1640培養基培養,并置于37 ℃、含5% CO2的細胞培養箱中。用Lipofectmine 2000分別將miR-NC、miR-224、anti-miR-NC、anti-miR-224、anti-miR-224聯合si-NC及anti-miR-224聯合si-ECRG4轉入A549細胞中,分別記為miR-NC組、miR-224組、anti-miR-NC組、anti-miR-224組、anti-miR-224+si-NC組和anti-miR-224+si-ECRG4組,培養48 h后,提取RNA和蛋白進行后續研究。
1.3.2熒光定量PCR檢測 根據RNAiso plus說明書提取NSCLC癌組織、癌旁組織標本和細胞系總RNA,采用PrimeScriptTMRT reagent Kit with gDNA Eraser試劑盒進行反轉錄,SYBR?Premix Ex TaqTM試劑盒檢測上述標本中miR-224和ECRG4的表達水平,分別以U6和β-actin作為內參。miR-224正向引物為5′-GAGCCAAGTCACTAGTGGT-3′,反向引物為5′-GTGCAGGGTCCGAGGT-3′;U6正向引物為5′-TGCGGGTGCTCGCTTCGGCAGC-3′,反向引物為5′-CCAGTGCAGGGTCCGAGGT-3′;ECRG4正向引物為5′-CCGGTTCTCCCTCGCAGCAC-3′,反向引物為5′-CGCTTCTGGCGCTTCAGGCT-3′;β-actin正向引物為5′-GAACCCCAAGGCCAACCGCGAGA-3′,反向引物為5′-TGACCCCGTCACCGGAGTCCATC-3′。每個細胞系設置3個復孔,重復檢測3次。
1.3.3Western blot檢測 用RIPA裂解液提取NSCLC癌組織、癌旁組織標本和細胞系總蛋白,采用BCA蛋白定量試劑盒檢測蛋白水平。采用SDS-PAGE電泳分離蛋白標本,并轉至PVDF膜上,5%脫脂牛奶封閉2 h,加入ECRG4和GAPDH抗體,置于搖床上4 ℃孵育過夜,然后用辣根過氧化物酶偶聯二抗室溫孵育2 h,ECL發光液孵育條帶并置于化學發光成像系統中進行曝光顯影,ImageJ軟件分析相應蛋白的相對表達水平。每個細胞系設置3個復孔,重復檢測3次。
1.3.4原位雜交和免疫組織化學檢測 采用5′和3′-雙地高辛標記的MIRURY LNA microRNA Detection Probe試劑盒進行原位雜交分析。采用VENTANA BenchMark XT染色系統對切片中的ECRG4蛋白進行孵育顯色。
1.3.5雙熒光素酶報告基因實驗 采用PCR擴增ECRG4的3′UTR,并將其克隆至pmirGLO報告質粒中構建pmirGLO-ECRG4-Wt重組質粒。采用TakaRa點突變試劑盒將pmirGLO-ECRG4-Wt中miR-224的結合位點進行突變,構建pmirGLO-ECRG4-Mut重組質粒。采用Lipofectmine 2000將miR-NC或miR-224分別與pmirGLO-ECRG4-Wt和pmirGLO-ECRG4-Mut共轉至A549細胞中,48 h后用Dual Luciferase?Reporter Assay System試劑盒檢測各轉染細胞熒光素酶活性。
1.3.6細胞增殖檢測 取對數生長期的各組細胞各3 000個接種于96孔板中,每組設置3個復孔,分別于24、48、72 h時每孔加入20 μL的5 mg/mL噻唑藍溶液,孵育4 h后棄去廢液,每孔加入150 μL 二甲基亞砜,然后用酶標儀震蕩10 min以溶解結晶,檢測490 nm處各孔吸光度值。
1.3.7細胞遷移、侵襲檢測 Transwell小室分為兩種,1種有預鋪基質膠用于檢測細胞侵襲,1種無預鋪基質膠用于檢測細胞遷移。取對數生長期的各組細胞,每組各取200 μL含1×105個細胞的細胞懸液加入Transwell小室上室中,下室中加入600 μL完全培養液,每組設置3個復孔。培養48 h后,經0.1%結晶紫染色后顯微鏡下觀察并拍照,計數各組遷移、侵襲細胞數。
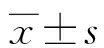
2 結 果
2.1NSCLC癌組織及癌旁組織中miR-224和ECRG4的表達水平比較 與癌旁組織相比,癌組織中miR-224表達水平明顯升高,差異有統計學意義(P<0.05),見圖1A、表1。癌組織中ECRG4 mRNA和ECRG4蛋白表達水平明顯低于癌旁組織,差異有統計學意義(P<0.05),圖1A、1B、表1。

注:A為原位雜交和免疫組織化學分別檢測NSCLC癌組織及癌旁組織中miR-224和ECRG4 mRNA(200×);B為 Western blot檢測NSCLC癌組織及癌旁組織中ECRG4蛋白。

表1 NSCLC癌組織和癌旁組織中miR-224和ECRG4的表達水平比較
2.2miR-224和ECRG4在各細胞系中的表達水平比較 與健康人肺支氣管上皮細胞系HBE相比,NSCLC細胞系L78、A549、H460中miR-224表達水平明顯升高,ECRG4 mRNA和ECRG4蛋白表達水平明顯降低,差異有統計學意義(P<0.05),見圖2、表2。

圖2 Western blot檢測細胞系HBE、L78、A549、H460中ECRG4蛋白
表2 細胞系HBE、L78、A549、H460中miR-224和ECRG4的表達水平比較
2.3ECRG4是miR-224的靶基因 利用microRNA.org網站預測發現,ECRG4可能是miR-224的潛在靶基因,見圖3。與miR-NC組A549細胞中miR-224的表達水平(1.00±0.08)比較,miR-224組A549細胞中miR-224的表達水平(4.15±0.46)明顯升高,差異有統計學意義(t=11.69,P<0.05)。miR-224和pmirGLO-ECRG4-Wt共同轉染的A549細胞中ECRG4-Wt熒光素酶活性(0.42±0.05)低于miR-NC和pmirGLO-ECRG4-Wt共同轉染的A549細胞(1.00±0.11),差異有統計學意義(t=8.31,P<0.05);而miR-224和pmirGLO-ECRG4-Mut共同轉染的A549細胞中ECRG4-Mut熒光素酶活性(1.03±0.09)與miR-NC和和pmirGLO-ECRG4-Mut共同轉染的A549細胞(0.96±0.10)比較,差異無統計學意義(t=0.90,P>0.05)。
2.4抑制miR-224可上調A549細胞中ECRG4蛋白表達 與anti-miR-NC組A549細胞中ECRG4蛋白表達水平(0.35±0.04)比較,anti-miR-224組A549細胞中ECRG4蛋白表達水平(0.96±0.09)明顯增加,差異有統計學意義(t=10.73,P<0.05)。anti-miR-224+si-ECRG4組A549細胞中ECRG4蛋白表達水平(0.13±0.01)明顯低于anti-miR-224+si-NC組(0.94±0.09),差異有統計學意義(t=15.49,P<0.05),見圖4。

圖3 預測的miR-224與ECRG4 3′UTR結合位點

圖4 抑制miR-224及聯合抑制ECRG4后A549細胞中ECRG4蛋白表達情況
2.5抑制miR-224對A549細胞增殖的影響 48 h和72 h時anti-miR-224組A549細胞吸光度值明顯低于anti-miR-NC組,差異有統計學意義(P<0.05);48 h和72 h時anti-miR-224+si-ECRG4組A549細胞吸光度值明顯高于anti-miR-224+si-NC組,差異有統計學意義(P<0.05),見表3。
2.6抑制miR-224對A549細胞遷移、侵襲的影響 anti-miR-224組遷移、侵襲細胞數均明顯低于anti-miR-NC組,差異有統計學意義(P<0.05);anti-miR-224+si-ECRG4組遷移、侵襲細胞數均明顯高于anti-miR-224+si-NC組,差異有統計學意義(P<0.05),見表4。
表3 抑制miR-224及聯合抑制ECRG4后A549細胞的吸光度值比較

表4 抑制miR-224及聯合抑制ECRG4后A549細胞遷移和侵襲細胞數個)
3 討 論
肺癌是全球腫瘤相關死亡的主要原因[11]。盡管近年來在手術和藥物治療方面取得了進展,但NSCLC患者的5年生存率仍然很低,其發病機制未完全闡明[12]。因此,深入研究NSCLC進展的分子機制將有助于發現潛在的治療靶標并優化現有的治療方法。
近年來,越來越多的研究集中于miRNA在腫瘤中的異常表達。相關研究表明,miRNA在大多數惡性腫瘤的發病機制中起重要作用,部分miRNA在腫瘤中作為原癌基因或抑癌基因發揮作用,并參與調控多種生物過程[13]。干擾miRNA的功能被認為是腫瘤治療的新策略。在NSCLC中,miR-326可通過靶向細胞周期蛋白D1抑制腫瘤進展,miR-22可通過靶向ATP檸檬酸裂解酶抑制骨肉瘤、前列腺癌、宮頸癌和肺癌的生長和轉移[14-15]。有研究發現,miR-224可通過靶向抑制SMAD4和人腫瘤壞死因子α誘導蛋白1促進NSCLC細胞生長和侵襲[11,16]。此外,miR-224在結直腸癌、肝細胞癌和腎癌等多種腫瘤類型中表達上調[17-19]。上述研究均表明,miR-224是一個促癌miRNA。本研究發現,miR-224在NSCLC癌組織和細胞中高表達,抑制其表達可抑制NSCLC細胞增殖、遷移和侵襲,與CUI等[11]和ZHU等[16]的研究結果一致。
ECRG4是NF-κB等主要生長因子信號通路的調節因子,可參與調控細胞周期進程、細胞遷移、衰老、祖細胞存活、分化及炎癥等[8]。大量研究發現,ECRG4在包括乳腺癌在內的多種腫瘤中表達下調或缺失[8,20]。有研究表明,低水平的ECRG4 mRNA可能與乳腺癌無病生存和遠處轉移的發生密切相關[21]。1項Meta分析表明,ECRG4 mRNA表達水平是乳腺癌無病生存率和總生存率的影響因素[22]。此外,在鼻咽癌中,ECRG4通常因啟動子過度甲基化而低表達,并且恢復其表達后可通過誘導自噬增強順鉑介導的細胞死亡[8]。上述研究表明,ECRG4可能作為一個抑癌基因在多種腫瘤中發揮作用。本研究發現,在NSCLC癌組織和細胞中ECRG4低表達,ECRG4是miR-224的靶基因,聯合抑制ECRG4可明顯逆轉抑制miR-224對NSCLC細胞A549細胞增殖、遷移和侵襲的抑制作用。
4 結 論
本研究證實,在NSCLC組織和細胞中miR-224高表達,ECRG4低表達,miR-224可通過靶向ECRG4來促進NSCLC A549細胞增殖、遷移和侵襲,進一步揭示了NSCLC發病的分子機制,可能為NSCLC的診斷和治療提供新的靶點。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
海峽科技與產業(2016年3期)2016-05-17 04:32:12
學苑創造·A版(2015年11期)2016-01-14 09:03:27
