維甲酸誘導N2A 細胞分化過程中CD2AP對突觸可塑性的調控
2020-08-24 13:26:44劉德山陳佃福張旭
中國現代藥物應用 2020年15期
劉德山 陳佃福 張旭
阿爾茨海默病(Alzheimer’s Disease,AD)是一種常見的、與老化密切相關的神經系統變性疾病,其發病率卻逐年升高[1],AD 預期的發病率將達到2001年的300%[2]。迄今已發現多個AD 的致病基因,如APP 基因、早老素基因1(PS1)和早老素基因 2(PS2)等,國際上多個研究團隊通過全基因組關聯分析(Genome-Wide Association Study,GWAS)發現了一系列散發性AD 的易感基因[3,4],前期研究提示CD2AP 表達水平下降可能參與AD 的發病[5]。目前國內外對CD2AP 在腎臟疾病中的致病機制研究較多,發現其參與多個信號通路并在維持細胞骨架中起著重要作用[6]。近期有國內外研究[7]以及本團隊前期的研究均提示CD2AP 參與AD的病理過程,但其在AD 發病中的具體機制仍不清楚。
突觸相關的結構組成主要包括Actin 細胞骨架、突觸后致密物質(postsynaptic dentisy,PSD)、谷氨酸受體等神經遞質受體和一些信號蛋白分子等。在研究中發現應用全反式維甲酸(all trans retinoic acid,ATRA)干預N2A 細胞后,發現CD2AP 的表達明顯升高,N2A 細胞神經突起明顯變長,轉染過表達CD2AP 后,神經突起顯著變長,而轉染shRNA 下調CD2AP 的表達水平后,神經突起顯著變短,提示CD2AP 在神經元骨架的形成中起著重要作用,提示CD2AP 可能在神經元突觸可塑性中起著重要作用。有研究報道神經元受損后在神經營養因子的作用下,CD2AP 可參與影響神經元骨架蛋白Actin 及受體的內吞作用。CD2AP 作為信號通路與細胞骨架重要的聯系分子,可與微絲骨架結合蛋白相互作用,在谷氨酸的誘導下,參與突觸形成、遷移等過程,本研究將通過體外ATRA 誘導N2A 細胞神經突起的生長,探討CD2AP 如何調節神經突觸的生長,為后續深入研究CD2AP 如何調控突觸可塑性提供重要理論依據。現報告如下。
1 材料與方法
1.1 實驗材料 ①細胞系:SH-SY5Y 細胞:購自中國科學院上海細胞所;N2A 細胞:本實驗室保種。②病毒:shRNACD2AP:山東維真公司;overexpressionCD2AP:上海吉凱公司。
1.2 方法
1.2.1 ATRA 干預N2A 細胞 復蘇N2A 細胞應用ATRA 干預(干預濃度為20 μmol/L,干預時間為5 d)后,密切觀察細胞狀態,繼續后續實驗。
1.2.2 病毒轉染 利用維真公司包裝的shRNACD2AP病毒和吉凱公司overexpressionCD2AP 病毒進行轉染;感染約72 h,觀察感染效率,進行后續實驗。
1.2.3 細胞免疫熒光 細胞爬片經處理后,放入12 孔板各孔中并于多聚賴氨酸進行包被。在轉染前1 d 鋪細胞(N2A),每孔鋪約1×105個細胞。第2 天轉染病毒或進行ATRA 干預、轉染或干預后進行細胞免疫熒光染色。
1.2.4 Western blot 從培養箱中取出細胞,抽提的總蛋白,采用Bradford 法測定蛋白質濃度,電泳、轉膜后用封閉液稀釋抗體[兔抗CD2AP單克隆抗體(1∶1000)、鼠抗GAPDH(1∶1000)、鼠抗β-actin(1∶1000)],孵育4℃過夜,用TBST 洗膜,再用封閉液稀釋相應的二抗,室溫下孵育1 h,TBST 洗膜后,采用Millipore 公司ECL化學發光試劑盒進行顯色,應用Image Lab 軟件進行化學發光成像、分析。
1.3 統計學方法 采用SPSS14.0 統計學軟件對研究數據進行統計分析。Western blot 數據以均數±標準差(±s)表示,進行Levene 方差齊性檢驗,方差齊時,組間兩兩比較采用LSD-t 檢驗;方差不齊時,采用Games-Howell 檢驗。P<0.05 表示差異具有統計學意義。
2 結果
ATRA 誘導N2A 細胞后,CD2AP 的表達明顯升高,N2A 細胞神經突起明顯變長;上調、下調N2A 細胞中的CD2AP 后,細胞突起顯著變長,ATRA 通過調控CD2AP 表達影響神經突起生長。
CD2AP 參與生長因子信號的蛋白復合物的支架,CD2AP mRNA 和蛋白在大腦中均有表達,Allen 腦圖譜表明CD2AP mRNA存在于特定區域的成年大鼠大腦中,特別是具有可塑性高的區域,為了確定CD2AP 是否參與對神經突觸生長的調節,在應用ATRA 進行干預(干預濃度為20 μmol/L,干預時間為5 d)的N2A 細胞中檢測了CD2AP 的表達情況。免疫熒光結果表明,N2A 細胞應用ATRA 干預后,其神經突起長度明顯變長。見圖1。同時,Western blot 法檢測結果表明ATRA 干預后,CD2AP 蛋白表達水平呈顯著升高。結果表明CD2AP蛋白水平受到ATRA 的正調控,并在N2A 細胞神經突起生長過程中起著重要作用。
為進一步研究CD2AP 對神經突起生長的調控,應用shRNACD2AP 和overexpressionCD2AP 兩種病毒分別對N2A 細胞進行上調和下調CD2AP 蛋白水平的干預。72 h 后,overexpressionCD2AP 轉染Western blot 檢測發現CD2AP 的表達量呈顯著升高,細胞神經突起長度明顯變長。見圖2。相反的,轉染shRNACD2AP 介導的CD2AP 表達減少導致了與蛋白質的減少程度一致的神經突起的長度顯著降低。見圖3。結果顯示,CD2AP 蛋白可能參與了神經突起生長的調控。
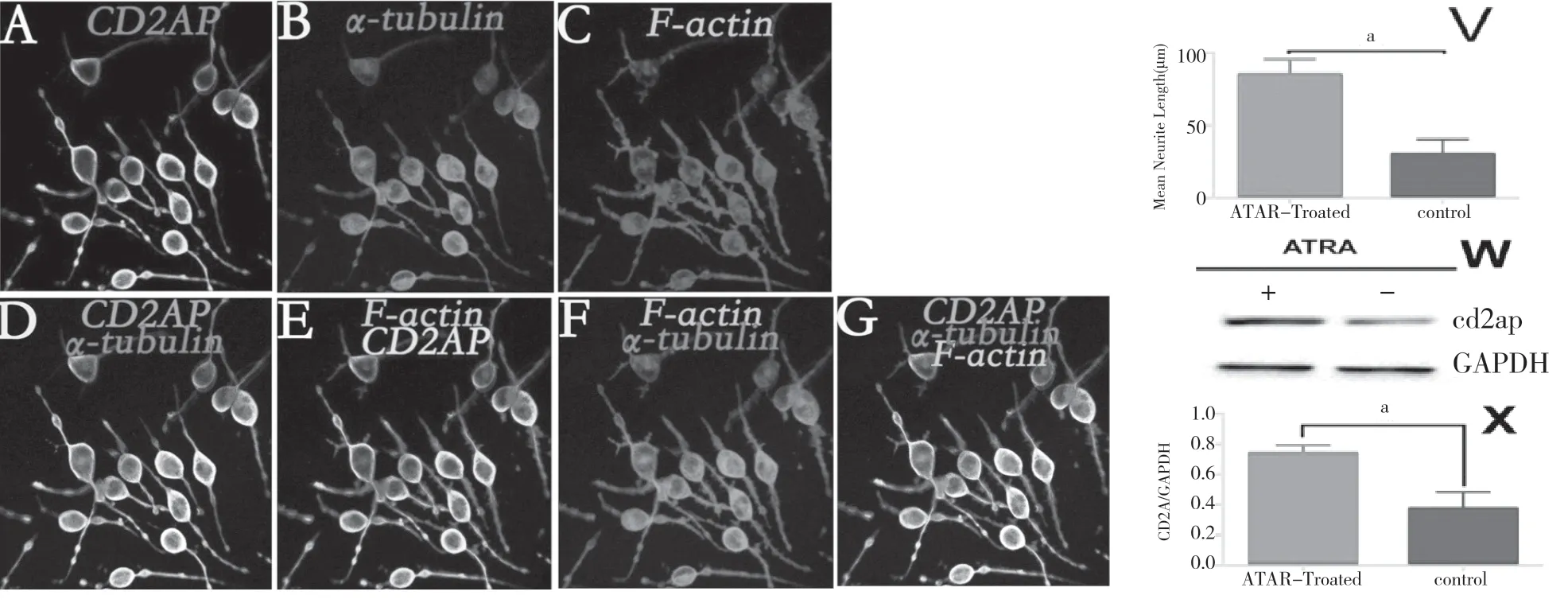
圖1 在N2A 細胞分化突起生長過程中CD2AP 表達水平顯著升高
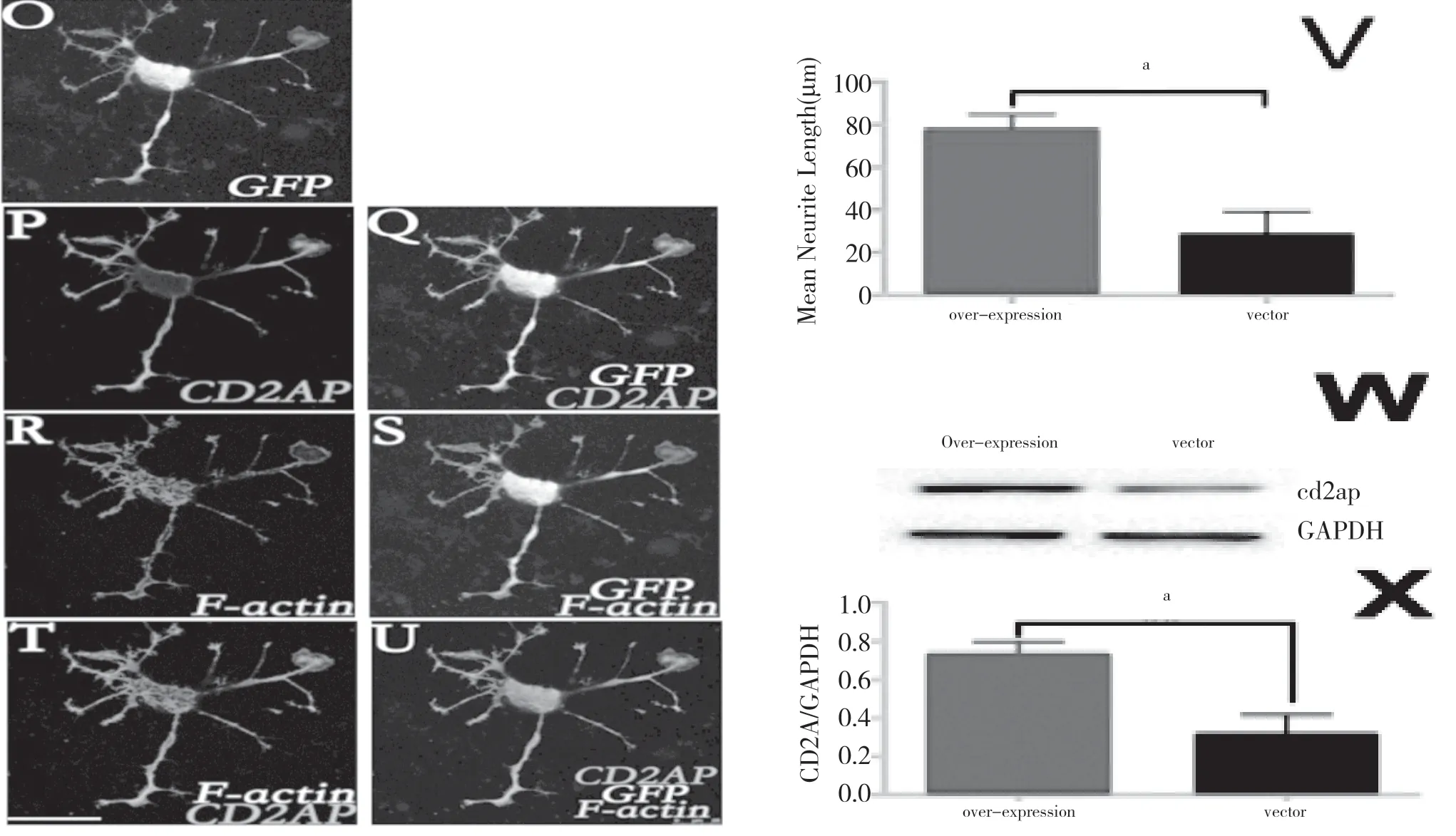
圖2 overexpressionCD2AP 干預N2A 細胞后CD2AP 表達水平顯著升高、神經突起顯著變長
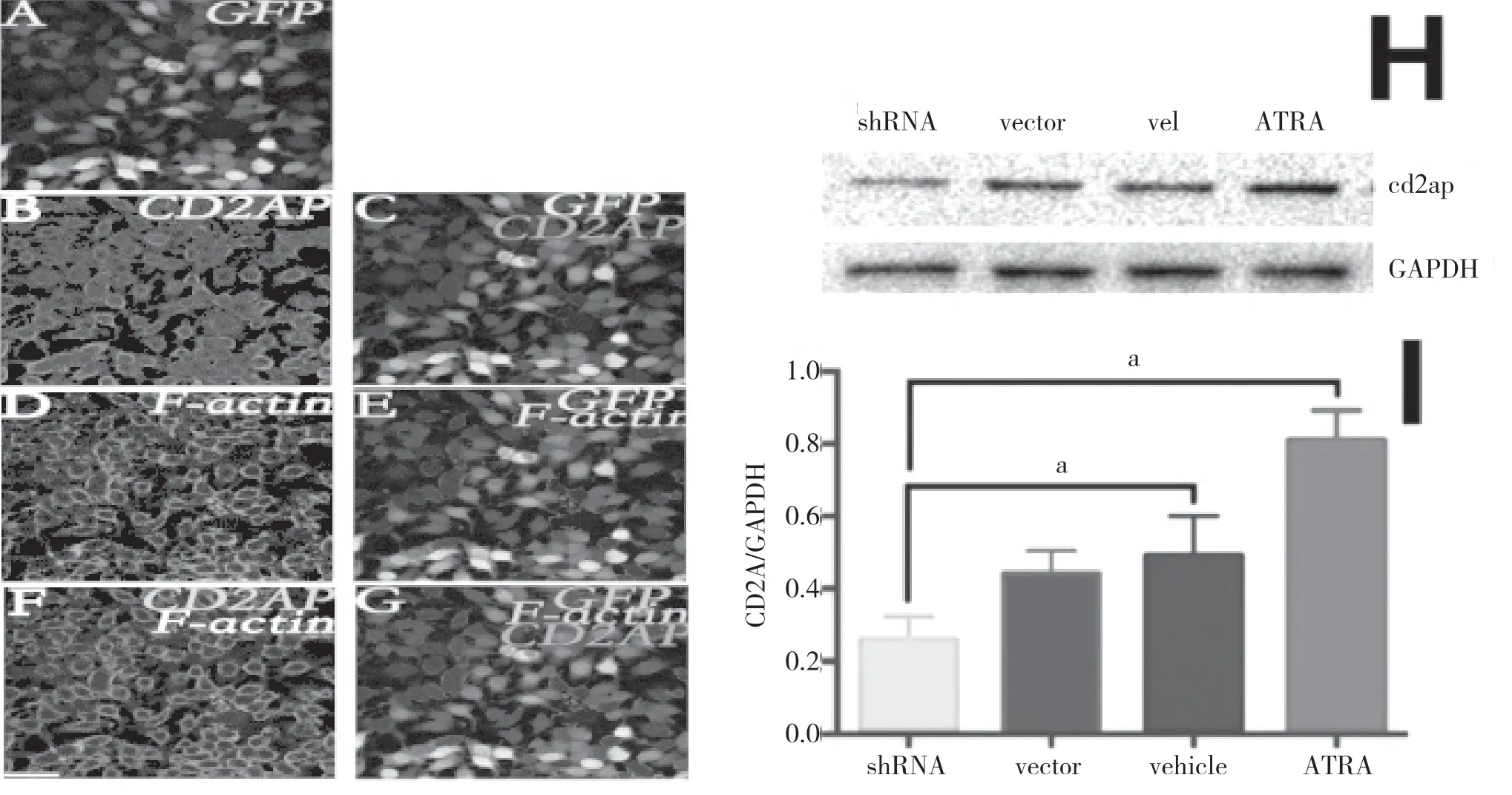
圖3 shRNACD2AP 干預N2A 細胞后,N2A 細胞CD2AP 表達下調、神經突起變短
3 討論
AD 其主要臨床表現為慢性、進行性認知功能損害,其起病隱襲,初始癥狀表現為記憶力缺失,癥狀會隨著病情進展逐漸加重,認知和記憶功能不斷惡化,AD 患者認知功能下降主要與突觸功能的異常相關,且突觸功能紊亂常常早于突觸的丟失。突觸前和突觸后相關蛋白均出現不同程度的下降,而且與認知功能的下降明顯相關,可見突觸功能的正常運轉以及維持與突觸相關蛋白的正常表達密不可分。突觸相關蛋白是突觸可塑性重要的分子基礎。有研究報道包括突觸結構和功能的變化是AD 患者和AD 轉基因小鼠模型腦部最早出現的表型,并且突觸水平的變化和認知功能損害的程度密切相關。而與突觸相關的結構組成主要包括Actin 細胞骨架、突觸后致密物質、谷氨酸受體[主要包括N-甲基-D-天冬氨酸受體(NMDAR)]等神經遞質受體和一些信號蛋白分子等。影響上述分子和信號通路的因素均可能影響突觸的結構和功能,進而可影響突觸可塑性。
在本研究中,應用ATRA 干預N2A 細胞后,發現CD2AP 的表達明顯升高,N2A 細胞神經突起明顯變長,轉染overexpressionCD2AP 病毒后,CD2AP 過表達,神經突起顯著變長,而轉染shRNA 下調CD2AP 的表達水平后,神經突起顯著變短,提示CD2AP 在神經元骨架的形成中起著重要作用。這和既往在腎病研究中報道的CD2AP 對調節足細胞的骨架結構起著關鍵作用的結果相一致[5],提示CD2AP 可能在神經元突觸可塑性中起著重要作用。
總之,ATRA 干預N2A 細胞后可上調CD2AP 的表達,或者overexpressionCD2AP 轉染細胞上調CD2AP的表達后,都可促進N2A 細胞神經突起的生長,而shRNACD2AP 下調CD2AP 的表達后,神經突起顯著變短,進而說明CD2AP 在神經突觸生長過程中起著重要的作用,目前有大量的研究表明AD 患者認知功能下降,其主要原因是突觸功能障礙[6,7]。這些結果對理解CD2AP 基因增加AD 易感性的特點提供了重要信息,從而為深入研究CD2AP 增加AD 易感性的發生機制獲得一點啟示,同時也為今后的研究提供了依據,在后續的研究中進一步深入探討CD2AP 調控突觸可塑性的信號通路。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
