瘦素對乳腺癌MDA-MB-231細胞MMP14調控作用的研究①
2020-07-07 06:15:50周雪清劉倩倩劉文慧楊姝婭駱耐香
華夏醫學 2020年1期
關鍵詞:乳腺癌
周雪清,劉倩倩,張 杰,劉文慧,楊姝婭,駱耐香③
(1.桂林醫學院免疫學教研室,廣西 桂林 541004;2.川北醫學院附屬醫院檢驗科,四川 南充 637000;3.桂林醫學院第二附屬醫院檢驗科,廣西 桂林 541199)
流行病學研究表明,乳腺癌已成為威脅全球女性健康和生命主要的因素之一[1],而肥胖與乳腺癌的患病風險和其臨床行為密切相關[2]。瘦素(leptin)是由脂肪組織分泌的與肥胖有關的重要激素,能夠與瘦素受體結合后通過調節飲食攝入和能量代謝參與體內內環境平衡的維持[3],同時它還參與了機體卵巢癌、胃腸道腫瘤等多種腫瘤的發生和發展[4]。膜型基質金屬蛋白酶(membrane-type matrix metalloproteinase-1,MT1-MMP)即MMP14屬于基質金屬蛋白酶家族,它直接或間接降解細胞外基質中的多種成分,與腫瘤的侵襲與遷移密切相關[5]。已知瘦素與MMP14均可促進乳腺癌的發生與發展[2-5],而兩者之間在乳腺癌是否有關聯目前尚無相關的研究報道。因此,本課題組以乳腺癌細胞系為研究對象,探討瘦素是否可以通過影響MMP14的表達促進乳腺癌發生和發展,為開發一種乳腺癌新型治療靶點提供思路。
1 材料與方法
1.1 細胞株與主要試劑
人乳腺癌細胞株MDA-MB-231購自中國科學院上海細胞庫。人重組瘦素購自美國 Peprotech公司,RPMI1640培養基及胰蛋白酶由美國Gibco公司提供,胎牛血清由美國Gemini公司提供,兔抗人MMP14抗體由Abcam公司提供,鼠抗人β-actin抗體及相關二抗由北京中山金橋生物有限公司提供,MTT試劑盒由北京索萊寶科技有限公司提供,ERK1/2、JNK、P38的信號通路抑制劑FR180204、SP600125、SB203580均由上海藍木化工有限公司提供。
1.2 細胞培養
人乳腺癌細胞株MDA-MB-231用含有10% 胎牛血清的RPMI1640完全培養基于37 ℃、5% CO2的細胞培養箱中溫育。
1.3 western blot法檢測細胞MMP14的蛋白表達水平
將處于對數生長期乳腺癌細胞接種于6孔板中,分對照組(不加瘦素)、不同濃度瘦素組(25,50,100,200,400 ng/ml)及瘦素+不同信號通路抑制劑組,作用24 h后提取各組細胞總蛋白,用western blot法檢測各組細胞MMP14的蛋白表達。實驗獨立重復3次。
1.4 MTT法檢測細胞增殖率
將處于對數生長期乳腺癌細胞經胰酶消化,制成單細胞懸液后,接種于96 孔板中,分二甲基亞砜(DMSO)組(溶劑組)、瘦素+DMSO組(對照組)及瘦素+不同通路抑制劑組,檢測作用0,1,2,3,4 d的吸光度值。每組設6個復孔,計算平均值。實驗獨立重復3次。
1.5 劃痕實驗檢測細胞遷移能力
取對數生長期乳腺癌細胞接種于12孔板中,分DMSO組、瘦素+DMSO組及瘦素+不同通路抑制劑組。瘦素作用前,用200 μl移液器槍頭在12孔板內垂直劃痕,PBS清洗2遍,然后換成含1%胎牛血清的培養基并做相應的組別處理。各組分別加入DMSO、瘦素+DMSO、瘦素+不同通路抑制劑(瘦素濃度100 ng/ml,各通路抑制劑濃度均為500 ng/ml),在鏡下觀察,根據乳腺癌細胞的生長周期選擇每12 h為1個觀察時間點觀察細胞遷移變化并拍照直至劃痕完全愈合,計算平均劃痕愈合率。實驗獨立重復3次。
1.6 統計學分析
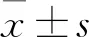
2 結果
2.1 不同濃度瘦素處理后乳腺癌細胞MMP14的蛋白表達水平
western blot結果顯示,與對照組相比,25,50 ng/ml瘦素組MMP14表達水平沒有差異, 100,200,400 ng/ml瘦素組MMP14表達水平明顯高于對照組(P<0.05或P<0.01),且隨著瘦素濃度的增加而升高。詳見圖1。

圖1 不同濃度瘦素處理乳腺癌細胞24 h后MMP14的蛋白表達(*P<0.05,P<0.01,*P<0.001)
2.2 瘦素聯合DMSO及不同通路抑制劑處理后乳腺癌細胞MMP14的蛋白表達水平
western blot結果顯示,與DMSO組相比,瘦素+DMSO組MMP14蛋白表達明顯升高(P<0.001)。與瘦素+DMSO組相比,瘦素+JNK 通路抑制劑組MMP14的蛋白表達量明顯降低(P<0.001),而瘦素+ERK組及瘦素+P38組MMP14蛋白的表達均明顯升高(P<0.01)。詳見圖2。

圖2 瘦素聯合DMSO及不同通路抑制劑處理乳腺癌細胞后MMP14的蛋白表達水平(P<0.01,*P<0.001)
2.3 瘦素聯合不同通路抑制劑處理后乳腺癌細胞增殖能力的變化
MTT結果顯示,隨著培養時間的延長,瘦素+DMSO組的乳腺癌細胞增殖率均高于DMSO組(P<0.05或P<0.01)。瘦素+不同通路抑制劑組的細胞增殖率在第1~2天與瘦素+DMSO組無明顯差異,在第3~4天則明顯低于瘦素+DMSO組(P<0.05或P<0.01),在第3天瘦素聯合JNK通路抑制劑組的細胞增殖率降低最為顯著。詳見圖3。




圖3 瘦素聯合不同通路抑制劑作用后乳腺癌細胞增殖情況(*P<0.05,P<0.01,*P<0.001)
2.4 瘦素聯合不同通路抑制劑處理后乳腺癌細胞遷移能力的變化
劃痕實驗結果顯示,瘦素+DMSO組的乳腺癌細胞遷移率在各檢測時間點均明顯高于DMSO組(P<0.05),瘦素+不同抑制劑組的細胞遷移率較瘦素+DMSO組均顯著降低(P<0.01)。詳見圖4。





圖4 瘦素聯合不同通路抑制劑作用后乳腺癌細胞遷移率(*P<0.05,P<0.01,*P<0.001)
3 討論
有研究表明,脂肪組織不僅在調節全身能量代謝中發揮重要作用,還能分泌包括瘦素和脂聯素等因子[6]。瘦素由位于7號染色體的OB基因編碼的蛋白質多肽類激素,主要由體內白色脂肪細胞合成分泌,包含167個氨基酸,分子量為16 KD[7]。瘦素通過與相應受體的結合,激活JAK-STAT通路以及絲裂原活化蛋白激酶(MAPK)、胰島素受體底物(IRS)、磷酸肌醇3激酶/蛋白激酶B(PI3K/Akt)、蛋白激酶C(PKC)和Rho家族GTPase等從而發揮作用[8-10]。體內瘦素的含量肥胖型乳腺癌患者高于非肥胖者,這與乳腺癌高侵襲性及不良預后關系密切[2]。瘦素可通過影響包括上皮間充質轉移、細胞與細胞外基質(ECM)的黏附以及ECM組分的蛋白水解等促進腫瘤轉移[11-12]。瘦素還可通過富集調節性T細胞,分泌免疫抑制因子,使腫瘤逃脫宿主的免疫監視,加速腫瘤的發生發展[13]。基質金屬蛋白酶(MMP)也在腫瘤發生和發展過程中促進腫瘤的侵襲和遷移[5]。已發現人類存在6種不同的膜型MMP(MT-MMPs),包括4種通過跨膜區域錨定在質膜上的Ⅰ型MT-MMP,即MT1-MMP、MT2-MMP、MT3-MMP和MT5-MMP,以及兩種糖基磷脂酰肌醇錨定的MT4-MMP和MT6-MMP。其中MT1-MMP(即MMP14)過表達可誘導乳腺癌的形成,促進乳腺癌的發生發展,其他小鼠上皮癌模型也發現MMP14的表達,特別是在TME的腫瘤相關細胞中,且均已證實其參與了腫瘤的進展[14]。MMP14缺失的乳腺癌小鼠模型顯示轉移減少,其作用機制是基質成纖維細胞降解Ⅰ型膠原減少[15]。
既然瘦素和MMP14均有促進乳腺癌的作用,本課題組選用MDA-MB-231乳腺癌細胞系,檢測瘦素對其MMP14表達的影響,探討其相關機制,進一步了解兩者在乳腺癌發生發展過程中的關系。研究結果顯示,低濃度瘦素對乳腺癌細胞MMP14表達無明顯影響,100 ng/ml及以上濃度均能顯著促進MMP14蛋白表達(P<0.05),且具有濃度依賴性。瘦素處理后,乳腺癌細胞的增殖和遷移能力均明顯升高,用ERK1/2、P38及JNK通路抑制劑分別和瘦素聯合作用細胞后,細胞的增殖和遷移能力均明顯降低(P<0.05)。用ERK1/2、P38通路抑制劑分別和瘦素聯合作用細胞后,均可顯著增加MMP14的表達水平(P<0.01),而JNK通路抑制劑和瘦素聯合作用細胞后則顯著降低MMP14的表達水平(P<0.001)。以上結果表明,瘦素可以促進乳腺癌細胞的增殖和遷移,可能通過ERK1/2、JNK及P38通路影響乳腺癌細胞MMP14的表達,其中JNK通路在失活狀態下可下調乳腺癌細胞MMP14的表達,而ERK1/2、P38通路在失活狀態下則上調MMP14的表達,促進乳腺癌細胞的增殖和遷移。有研究報道,瘦素還通過調節細胞周期蛋白D1、IGF-I、E-cadherin、血管內皮生長因子及其受體2、P53、Survivin以及多種組織因子的表達,調節乳腺癌增殖、黏附、侵襲、遷移、炎癥和血管生成等多種重要分子的作用[3]。本研究尚有不足,例如,細胞系單一、未涉及瘦素受體的相關實驗以及基因水平,有待于進一步研究。
本研究表明,瘦素可通過對MMP14蛋白表達的調控促進乳腺癌細胞的增殖和遷移。瘦素調控MMP14的作用可通過對ERK1/2、JNK及P38這3條信號通路的關鍵信號分子調控來實現。本研究創新性地提出了瘦素與MMP14的相關性,為開發新型乳腺癌診斷與治療靶點提供實驗依據。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
現代臨床醫學(2022年1期)2022-02-12 02:04:58
甘肅科技(2020年20期)2020-04-13 00:30:42
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
幸福家庭(2019年14期)2019-01-06 09:15:38
祝您健康(2018年5期)2018-05-16 17:10:16
癌癥進展(2016年9期)2016-08-22 11:33:20
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:08
