西北太平洋低緯度區域海水中溶解氨基酸的分布及組成研究
2020-06-03 09:21:40高先池楊桂朋
海洋科學 2020年5期
關鍵詞:區域
張 寧, 陳 巖, 高先池, 楊桂朋,
(1. 中國海洋大學化學化工學院, 山東 青島 266100; 2. 青島海洋科學與技術試點國家實驗室海洋生態與環境科學功能實驗室, 山東 青島 266237)
氨基酸(AAS)遍布海水、顆粒物和沉積物中, 是生物體有機氮(ON)和有機碳(OC)的主要存在形式之一[1], 其合成、降解過程與海洋生物緊密聯系, 是自養生物的重要氮源和異養生物的重要碳、氮源, 也是海水中OC和ON的重要儲庫。作為海洋中半不穩定有機物(semi-labile dissolved organic matter, SLDOM)的代表之一[2-3], AAS是海水中最先被異養生物礦化為二氧化碳的物質[4], AAS的降解會顯著降低溶解有機質(DOM)對異養生物的營養價值[5]。溶解氨基酸(THAA)占溶解有機氮(DON)含量的10%左右[6-7], 它的濃度變化受水體中生物代謝、海水運動、水團混合等多種因素的影響。高循環速度、與生物活動高度關聯是 THAA的特點, 也是前人將它作為溶解有機物(DOM)的生物可利用性指示劑的原因[8-9], 之前的研究表明, 異養細菌可選擇性降解不同種類的AAS, 故可利用 THAA組成變化指示海水中有機質的生物可利用性和降解程度[8]。因此研究海水中THAA的分布、組成及其與環境因子的相關性, 可以反映有機質的降解情況, 對于揭示相應海區有機碳、氮的生物地球化學行為有著重要的意義。
THAA既易受生物活動影響, 又是溶解有機物的重要組成部分, 因此AAS的碳歸一化產率(THAA-C%)被廣泛用于海洋有機質早期成巖狀況的定量描述[10]。隨著有機物降解程度的增加, THAA-C%呈降低趨勢。THAA-C%此特點可以指示有機物初期的降解狀態[11],它可指示有機物在降解過程中的量變。Davis等通過培養實驗證明THAA-C%可在20 ~ 30 d內降至初始值的50%, 并認為此參數可以準確表征DOM在降解初期的特征[11]。在THAA的降解過程中, 14種AAS并未表現出協同性, 而是會被選擇性的去除或富集, 并且 AAS的來源不同也會導致其組成差異,Dauwe 和 Middelburg發現此現象后運用主成分分析法(PCA)總結出降解因子(degradation index: DI), 從組成變化角度入手描述這種差異[8]。DI值范圍通常為-1~1.5, 越接近范圍下限代表DOM被降解的程度越大, 樣品中AAS可能來源于陸源或深海DOM; 越接近1則代表樣品遭受降解的程度越輕, 組成越接近海源現場生產的 DOM[8,12]。相對于常用于在 DOM 降解初期使用的 THAA-C%,DOM 降解的中期更宜使用 DI值表征[13-14], 是指示THAA降解過程中組成變化的參數。
西北太平洋低緯度海域的物質交換與能量交換強度非常高, 是研究熱帶海域中物質與環境關系的典型區域[15-17], 此處水團和環流是區域內能量交換的重要載體, 也是水體分配的主要推動者[18-20]。國際上對溶解態氨基酸的研究開展較早[21-23], 而國內對河流及近岸海域海水中 THAA的調查也在不斷增加[24-26], 但關于西北太平洋低緯度區域的 THAA的研究截至目前尚未被廣泛報道。本文選取西北太平洋低緯度海域為研究對象, 調查了該區域中 THAA的濃度分布及組成, 探討了 THAA與溶解有機碳(DOC)、葉綠素(Chl a)、海流等環境因子的關系, 并運用基于氨基酸組成的降解因子DI和THAA-C%對該海域有機質的降解情況進行了分析。
1 材料和方法
1.1 采樣區域
于2018年10月5號至11月6號搭乘“科學號”對西北太平洋低緯度區域上層海洋(5~200 m)進行取樣, 本文取樣層為5層, 分別為5 m、50 m、葉綠素最大層(Subsurface chlorophyll maximum, SCM, 深度約為100 m)、150 m、200 m。圖1(a)顯示出本次航次取樣的兩個斷面24個站位。圖1(b)展示出西北太平洋低緯度區域表層的基本環流結構。根據前人文獻將 E系列站位中的 2°N—5°N劃為 HE區, 5°N—7°N 為 NECC 區, 7°N—9°N 為 ME 區, 9°N—14°N 為MC區, 14°N—21°N為NEC區, 此外, 圖中未標注的南赤道流在0°N—5°N處也有影響。
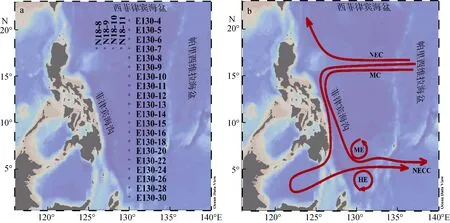
圖1 2018年秋季西北太平洋低緯度區域調查站位(a)、海流(b)示意圖Fig. 1 Schematic of survey stations (a) and currents (b) in the low-latitude area of the Northwest Pacific in the fall of 2018
1.2 樣品采集和保存
海水采集通過10 L Niskin Rosette 采水器進行采集, 水樣采集后立刻使用Whatman GF/F濾膜進行低壓過濾, 濾液轉移至硼硅玻璃瓶(450℃高溫灼燒4 h)中, 濾膜放于用鋁箔紙包裹的離心管中, 濾液、濾膜置于-20℃環境冷凍保存。溫度、鹽度數據由CTD現場測得。GF/F濾膜、樣品瓶、鋁箔等均在450℃條件下高溫灼燒5 h以除去有機碳。
1.3 樣品分析
1.3.1 THAA的測定
樣品采用實驗室已建立的鄰苯二甲醛-巰基丙酸(OPA-MPA)柱前衍生高效液相色譜法測定[27]。流動相: A是0.05 mol/L的乙酸鈉緩沖溶液(pH = 7.2 ± 0.05),且乙酸鈉溶液∶四氫呋喃 = 99∶1; B為甲醇∶乙腈∶水(v/v/v)=1∶1∶1。流動相均需通過 0.2 μm (PALL Supor-450)聚醚砜濾膜除去顆粒物。色譜柱為Agilent ZORBAX Eclipse 氨基酸分析柱, 測樣時使用梯度分離。氨基酸標準圖如圖2。
取2 mL海水樣品與2 mL優級純濃鹽酸(12 mol/L)混合于5 mL安瓿瓶, 加入40 μL 20% (w/v)抗壞血酸,通入高純氮氣1 min除去空氣, 密封后, 在110 ℃環境下高溫水解 22 h。水解完畢后取 2 mL水解液在60℃下進行真空旋轉蒸發旋蒸, 向蒸干后的樣品中加入1 mL Milli-Q水, 超聲振蕩2 min使樣品再溶解。取再溶解的溶液760 μL和衍生化試劑(3-MPA溶液)240 μL混合后避光衍生化19 min后, 進樣測定,測得濃度即為樣品中THAA濃度。

圖2 氨基酸標準的液相色譜圖 (0.5 μmol/L)Fig. 2 Chromatogram of the amino acid standards (0.5 μmol/L)
1.3.2 DOC的測定
海水樣品使用Whatman GF/F玻璃纖維濾膜(d =47 mm)進行低壓過濾, 轉移至硼硅玻璃瓶中-20℃冷凍避光保存。帶回實驗室后使用島津總有機碳分析儀 TOC-VCPH測定 DOC, 選用的催化劑和分析方法分別為 Pt催化劑、高溫催化氧化法。DOC樣品在680℃下平行進樣3~5次后取其平均濃度, 該方法測定的相對標準偏差 < 2%。儀器空白經邁阿密大學Hansell實驗室低碳水校正。
1.3.3 Chla的測定
定量過濾300 mL海水樣品, 濾膜快速置入15 mL離心管內-20℃冷凍保存。測定時在存放有膜樣的離心管中加入 90% (v/v)的丙酮水溶液, 避光冷藏萃取24 h, 使用SC-3610離心機進行低速離心, 離心后取離心后的上清液使用日立F-4500熒光分光光度計測定, 測定前需使用Jeffrey-Humphrey方程式對二級母液進行校正, 該方法的最低檢出限為0.01 μg/L。
1.3.4 DIN的測定
將海水樣品通過 Whatman GF/F玻璃纖維濾膜(d = 47 mm)進行低壓過濾后轉移至聚乙烯瓶中, 在-20℃條件下冷凍保存。帶回實驗室后, 立即使用Seal Analytical AA3營養鹽自動分析儀進行分析, 測得-N、-N、-N, DIN為- N、N、N三種形態N的總和。檢出限0.04 μmol/L,檢出限 0.003 μmol/L,檢出限0.02 μmol/L; 測定精密度小于3%。
2 結果和討論
2.1 西北太平洋低緯度區域海水中THAA、Chl a、DOC的水平分布特征
各站位海水中THAA、Chla、DOC的濃度范圍列于表1。
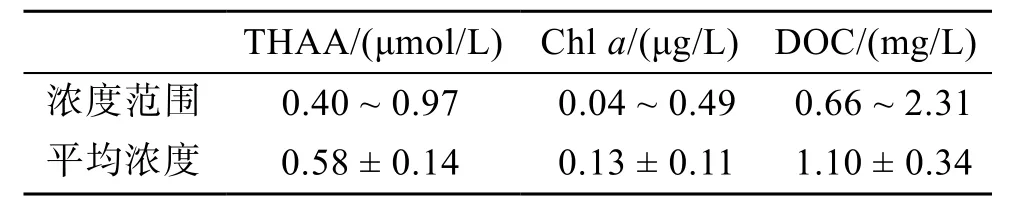
表 1 西北太平洋低緯度區域表層海水中(5 m)THAA、Chl a、DOC的濃度范圍Tab. 1 Concentration range of THAA, Chl a, and DOC of surface seawater (5 m) in the low-latitude area of the Northwest Pacific
在該調查海域, 表層海水(5 m)中THAA、Chla、DOC的濃度范圍列于表1, 該海域THAA濃度結果明顯低于春季北黃海表層海水[25]、南海北部表層海水[28]、東海表層海水[29-30], 高于同處西北太平洋的日本伊勢灣海域的表層濃度[31]。表層THAA濃度從南至北呈現出低-高-低-高的趨勢, 整體濃度低于中國近海[19,28-30]。THAA高值區出現在NEC區和ME區,NEC分叉點隨深度的降低向南偏移, 分叉點年平均位置在14°N且在10月份會北移[32-33], 此處高值區的出現原因可能是因為自東向西的NEC攜帶有高濃度的THAA向上擴散導致的; DI值和THAA-C%結果表明, ME高值區處降解程度較大, 且此處未存在Chla高值但存在DOC高值, 推測此處高值并非是浮游植物現場生產, 而是由于陸源輸入所致; 2°N—6°N 是THAA低值區, 而Chla在此處出現高值, 后文DI、THAA-C%表明該區域有機物的降解程度很高, 可能是由于此處溫度、光照條件促進了THAA的細菌降解和光降解, 另一個原因可能與ME上升流有關, 前人證明此處存在 Ekman抽吸引起的上升流[34], 上升流將下層低濃度、高降解程度的THAA帶到表層也是產生此現象的原因之一。此次航次測定Chla的濃度低于中國近海[25,28-30,35]的Chla濃度, 與齊雨藻報道的西太平洋濃度近似[36]。Chla的濃度從南到北呈下降趨勢, 出現此現象可能是因為中部和北部的寡營養鹽狀態是此處浮游植物生長的主要限制因素,而 NECC攜帶的大量營養鹽支持了南部浮游植物的生長; DOC的水平分布不均勻, 在9°N出現濃度最大值, 11°N 出現濃度最小值。在 1°N—9°N 和 11°N—21°N 均呈現為升高趨勢, 且 9°N 以南的平均濃度(1.24 ± 0.44 μmol/L)大于 11°N 以北的平均濃度(1.02± 0.18 μmol/L)。在 ME 高值區存在 THAA 高值, 而在此區域并未出現Chl a、DIN、T、S等參數的高值, 猜測此處DOC、THAA的高值可能是亞洲風塵帶入所致[37-38]。
將THAA、DIN、Chl a、DOC等參數進行相關性分析, 結果顯示均不存在顯著相關性。這可能是因為 THAA作為 SLDOM的重要組分, 其產生、循環與降解會受到浮游生物、光照、溫度、DO、水團運動等諸多因素的影響。此海域氨基酸碳/溶解有機碳(即 THAA-C%)較低, THAA的濃度降低并不能引起DOC濃度的顯著變化, 這可能是 THAA與 DOC未呈現顯著相關性的原因之一。結合該海區內海流復雜、初級生產力低、光照強烈的具體情形, 各環境因子對 THAA施加的影響可能是多因素協同作用, 因而THAA的濃度與單一參數不呈顯著相關。

圖3 西北太平洋低緯度區域表層海水中THAA(a)、chl a(b)、DOC(c)的水平分布Fig. 3 Horizontal distributions of THAA (a), Chl a (b), and DOC (c) in surface seawater of the low-latitude Northwest Pacific
2.2 西北太平洋低緯度區域 130°E斷面上層海洋中THAA、Chl a、DOC的垂直分布特征
為了探究各個環流對西北太平洋低緯度區域上層海水中 THAA 分布的影響, 我們設置了在 130°E上2°—21°N區域內的10個站位進行垂直(5 ~ 200 m)方向的采樣調查。此處區域是海流密集區, THAA、DOC、Chl a的分布受到了海流的強烈影響。E斷面THAA、Chl a、DOC的垂直濃度分布列于表2。
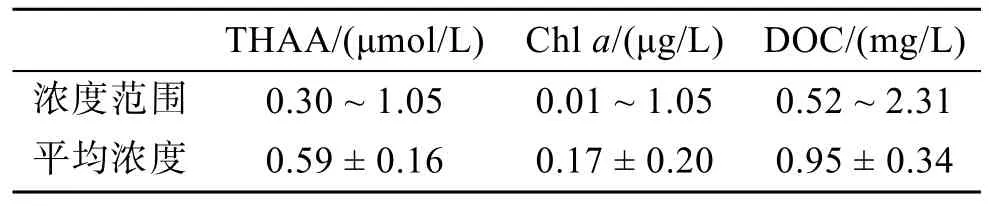
表 2 西北太平洋低緯度區域垂直斷面(E)中 THAA、Chl a、DOC的濃度范圍Tab. 2 THAA, Chl a, and DOC concentration ranges at vertical section (E) of the low-latitude Northwest Pacific
圖4(a)為該斷面溫度的垂直分布圖, 由圖可知在ME區存在上升流, 且該水域的水體溫度分層明顯,圖 4(b)中可看出該海域為高鹽度海域, 鹽度分布平均,表層的鹽度略低于下方水體。由表 2和圖 4(c)可知,該海域 THAA濃度較低, 但高于同為低生產力的楚科奇海[39], 介于西北太平洋與伊勢灣濃度之間[31], 此處低 Chla濃度反映出低初級生產力, 這與 THAA、DOC整體濃度較低相吻合。在圖4(a, c)中可以看到THAA 的低值區和 ME 上升流區重合(0°N—9°N,50 ~ 200 m深處), 該處THAA-C%低于臨近區域, 表明此處有機質已遭受了較長時間的初期降解, 由于較低的水溫更適宜異養生物生長[40-42], 強烈的細菌降解作用可能是此處THAA主要的去除途徑。THAA的濃度在 9°N—21°N真光層內隨深度增加而逐漸增加, 表層的高溫和寡營養鹽狀態共同限制了浮游植物的生長[36-37], 浮游植物生長受限、光降解[43-44]與細菌降解三者共同作用使表層THAA呈現低濃度。而在表層水以下, THAA濃度轉高, 這可能是由于上層浮游植物死亡后的沉降過程會發生自溶和異養細菌分解作用轉化為 THAA[29], 且溶解氧濃度隨深度增加而降低, 這也會限制THAA的細菌降解[45]。Chla濃度在5 ~ 200 m變化很小, 只在100 m深度附近出現SCM, 該現象出現原因是因為在此深度DIN高于表層, 且大洋區水體透光度高, 能支持熱帶西太平洋海區適應高鹽環境的浮游植物生長[36]。表層濃度低主要是因為開闊大樣區域表層寡營養鹽且水溫高,浮游植物生長受限[37], 而下層 DIN濃度雖較高但缺乏光照, 光限制了浮游植物的生長, 導致Chla濃度較低; Chla的高值區出現在5°N的50 m深, 可能是由于此處的ME伴隨的上升流將底部DIN帶到該深度, 強烈的光照使得浮游植物旺盛生長所致。DOC濃度整體呈現出隨深度增加而降低的趨勢, 其高值區出現在10°N附近的0 ~ 50 m, 可能是受到陸風輸送的影響[37-38]。
將上層海洋中的THAA、DOC、Chla濃度進行相關性分析, 發現三個參數間并未表現出顯著相關性, 這可能是因為三種參數既與生物活動息息相關,又易受到水團輸送、光照和陸源輸送的影響, 加之此海域海流、水團紛繁復雜互相影響, 導致參數之間互不表現出相關性。
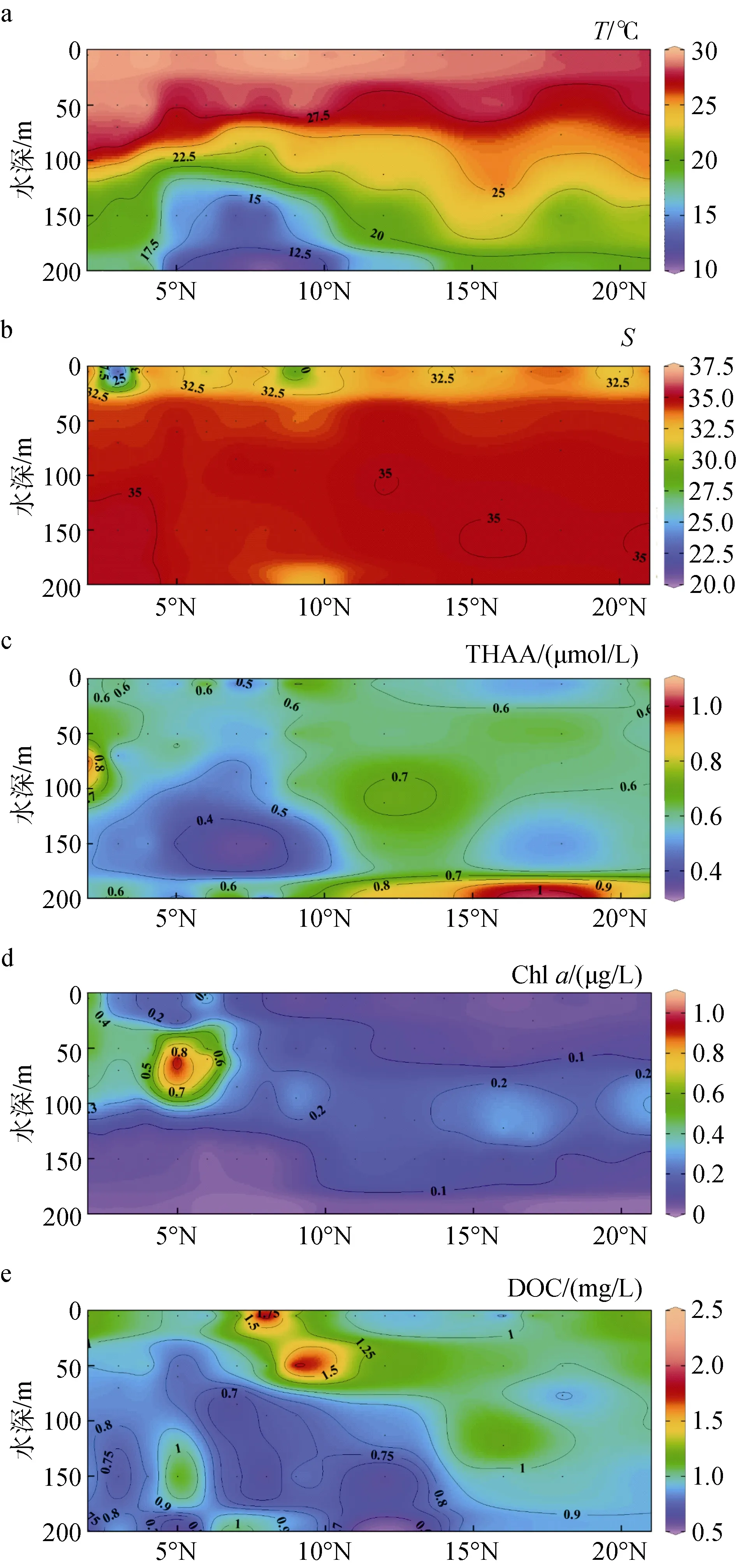
圖4 西北太平洋低緯度區域130°E斷面上層海洋中水溫(a)、鹽度 (b)、THAA (c)、Chl a (d)、DOC (e)的垂直分布Fig. 4 Vertical distribution of temperature (a), salinity (b),THAA (c), Chl a (d), and DOC (e) concentrations in the upper ocean on the 130°E Section of the lowlatitude Northwest Pacific
2.3 西北太平洋低緯度區域海水中 THAA的組成
THAA根據側鏈基團的不同可分為酸性、堿性、中性、芳香類、含羥基類、含硫氨基酸六類。西北太平洋低緯度區域海水中溶解氨基酸中各單體 AAS的相對組成如圖5所示。
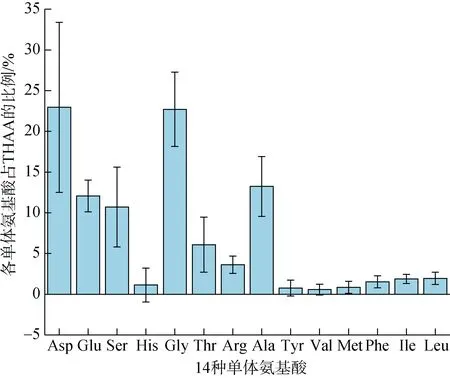
圖5 單體氨基酸占THAA的摩爾百分比Fig. 5 Monomeric amino acids as a percentage of THAA
測定的14種AAS占比差異較大, 6種優勢AAS(Asp、Glu、Ser、Gly、Thr、Ala)的總和可占到 THAA的87.74%, 其中Asp相對占比最高, 可達THAA的22.95%, 這與之前所報道的亞得里亞海[46]的結果相似, 說明在海水體系中 THAA的組成近似。根據董良[47]對西太平洋的研究, 西太平洋海域優勢藻類是硅藻, 硅藻細胞的主要構成成分是 Asp、Ser和 Glu等[48], 浮游藻類死亡后的細胞破裂和被浮游動物攝食會將體內的溶解氨基酸釋放到周圍海水中, 成為THAA的重要來源之一。
各類AAS的平均摩爾百分比如圖6所示。它們所占比例大小依次為中性氨基酸 > 酸性氨基酸 >含羥基類氨基酸 > 堿性氨基酸 > 芳香類氨基酸 >含硫氨基酸, 在此處海域中, 中性氨基酸的平均摩爾百分比略高于酸性氨基酸, 這兩種氨基酸共占所測14種溶解氨基酸的75.34%, 在此海域中是優勢氨基酸。
基于THAA的標準化數據(各單體AAS占THAA的摩爾百分比)做相關性矩陣, 如表 3所示。源自此次的數據顯示THAA中有12對氨基酸呈顯著負相關(Asp-Ser, Asp-His, Asp-Gly, Asp-Thr, Asp-Tyr, Asp-Ile,Asp-Leu, Glu-Ala, Glu-Leu, His-Gly, His-Ala, Gly-Tyr), 說明這12對個體AAS在海水中的動態行為有所差異; 同時有 15對 AAS呈顯著正相關(Ser-Gly,Ser-Thr, Ser-Leu, His-Tyr, His-Thr, Gly-Thr, Gly-Leu,Thr-Tyr, Tyr-Met, Val-Met, Phe-Ile, Phe-Leu, Met-Ile,Met-Leu, Ile-Leu), 這說明15對單體AAS在海水中具有相似的生物地球化學行為。
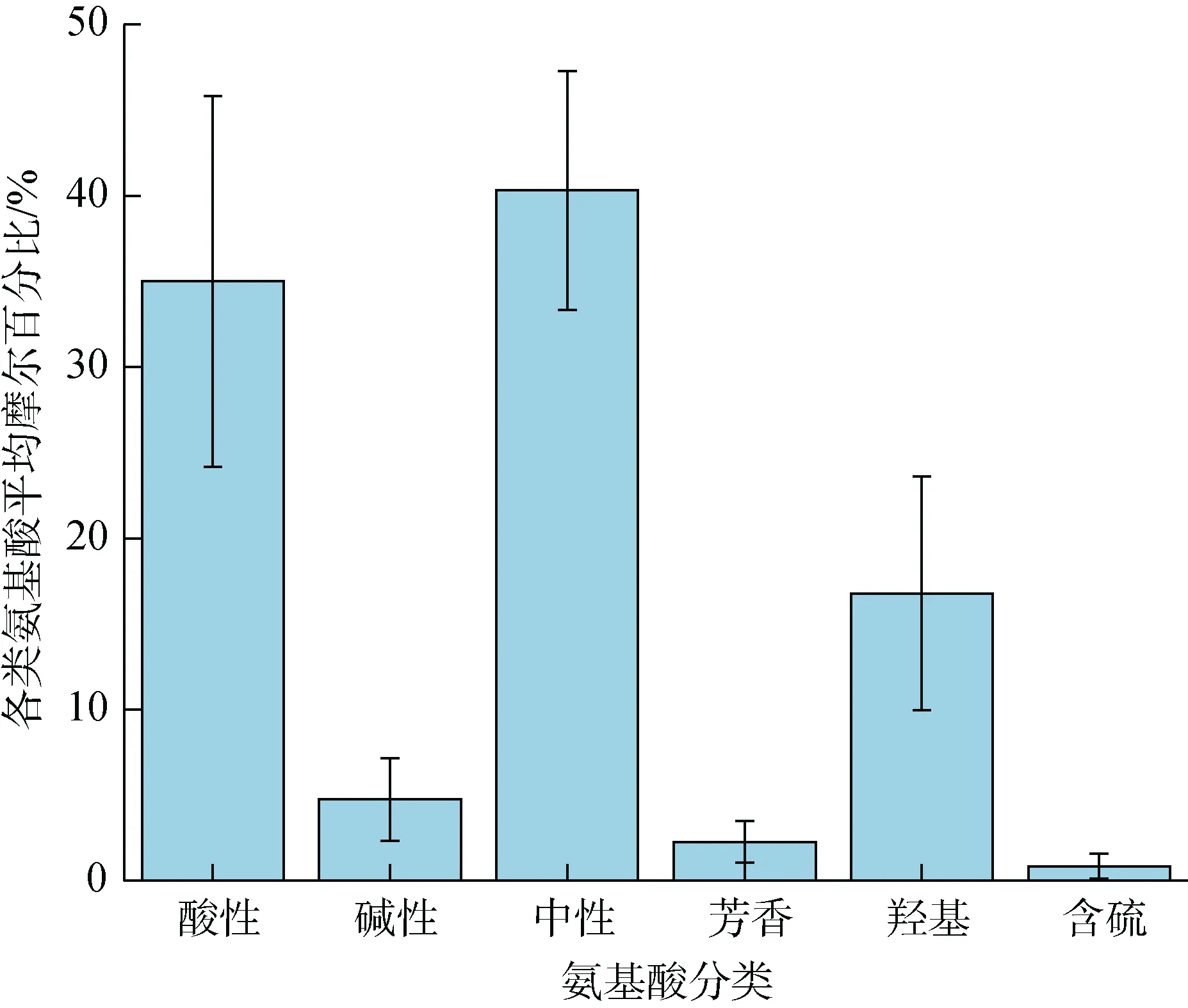
圖6 THAA中酸性、堿性、中性、芳香和含硫氨基酸的平均摩爾百分比Fig. 6 Average mole percent of acidic, basic, neutral, aromatic,and sulfur-containing amino acids in THAA
2.4 利用THAA-C%和DI值指示該海域有機物降解狀態
圖7 顯示了DI值和THAA-C%在西北太平洋低緯度區域表層海水的水平分布, 計算得出 THAAC%值的范圍是 0.89% ~ 3.75%, 平均值為 2.25% ±0.61%。相對于浮游植物體中24% ~ 49%的THAA-C%范圍[49], 該海域的2.25% ± 0.61%的均值證明此處了初級生產力極低。浮游植物釋放出的THAA進入表層海水后, 經歷高強度、長時間的光降解與細菌降解[11,49],而低生產力制約了新鮮 THAA的補充, 使表層海水中產生了DOM高降解程度的現象。在此區域內DI值的范圍為-2.27 ~ -1.15, 均值為-1.83 ± 0.29, 顯著低于中國近海和日本伊勢灣, DI與THAA-C%共同證明此區域內的氨基酸已經歷了較長時間的降解, 年齡較老。
DI、THAA-C%在該海域的垂直分布如圖8所示,其中THAA-C%的范圍是0.89% ~ 7.04%, 平均值為2.80% ± 1.05%, 該范圍高于楚科奇海[39]、西北太平洋和日本伊勢灣[31], 和表層 THAA-C%進行對比發現,50 ~ 200 m深度的THAA相對更“新鮮”, 降解程度低于表層THAA, 由于Chla出現了典型熱帶性次表層葉綠素最大值, 猜測是次表層生產力更高和光降解程度低于表層所致。在此研究中, THAA-C%的高值區出現在 10°N—21°N(3.32% ± 1.32%, n=15), 低值區出現在 2°N—9°N(2.56% ± 0.78%, n=33), 這個值與日本伊勢灣[31]、其他海洋環境[50]中報道的值十分相似。圖8(a)顯示了該區域內DI值的垂直分布, DI值范圍為-2.27 ~ 2.82, 平均值為-1.56 ± 0.74, 低于DI值-1 ~ 1.5的常見范圍, 推測該區域的THAA主要來自降解程度較高的物質。DI在5 ~ 200 m深度內隨深度增加呈上升趨勢, 與 THAA-C%體現出的趨勢相同, 這個趨勢與以往研究相反, 這可能是由于200 m對大洋而言僅為真光層, 在透光度高的赤道太平洋中, 上層數十米由于光照強烈加速了 THAA的光降解[43-44]、細菌降解[37], 隨著深度增加光照強度隨之減弱, THAA的光降解受到限制, 導致基于THAA的降解因子反映出上層降解程度高于下層的趨勢。據前人的研究, 海洋浮游植物的直接釋放并非THAA來源的主要因素[51], 上層浮游植物死亡后的
沉降過程會發生自溶和異養細菌對浮游植物殘骸的分解[29], 這兩種作用會將顆粒態AAS轉化為THAA而增加下方水體中 THAA的濃度, 而下方水體中光降解與細菌降解強度由于光照和 DO等條件的限制要低于上方水體[37,43-44], 導致SCM未與THAA-C%、DI的高值區共同出現。
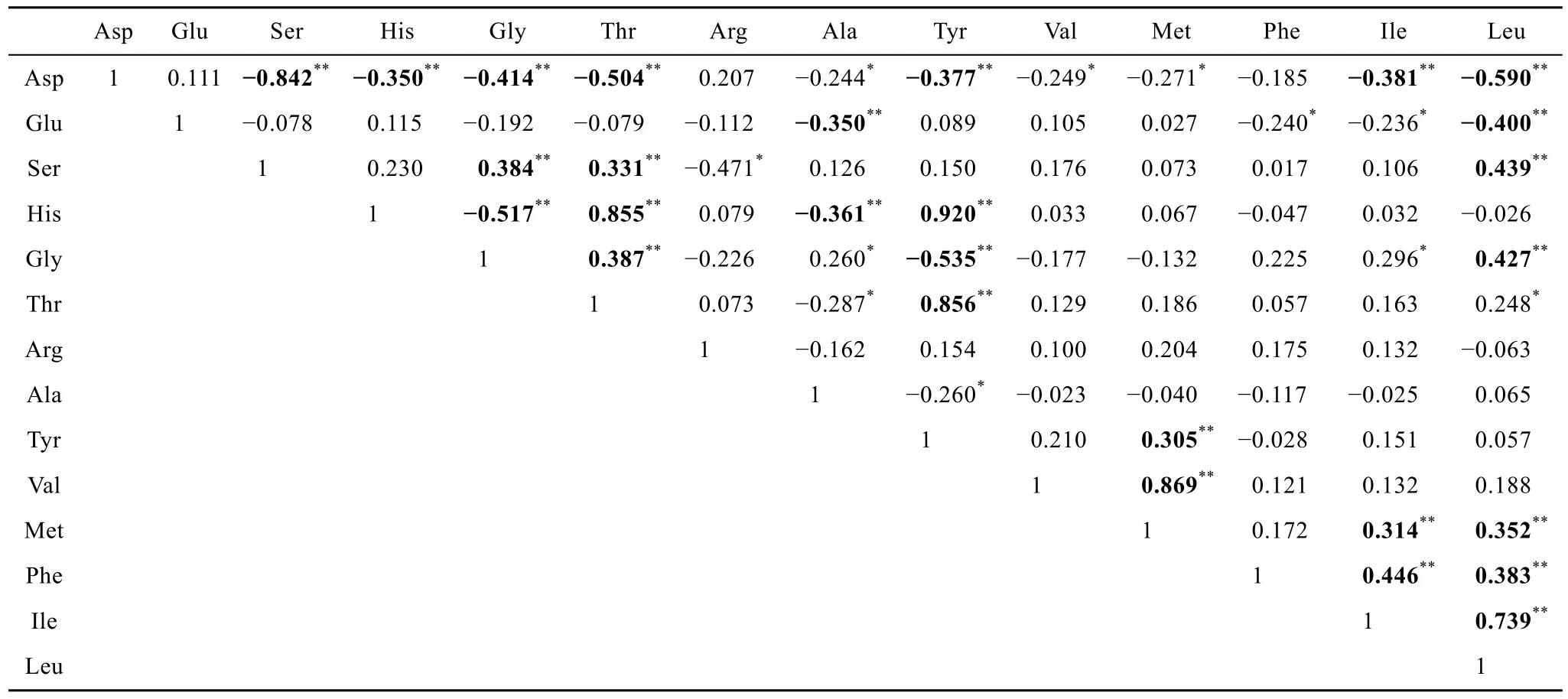
表3 西北太平洋低緯度區域海水中THAA單體氨基酸相對含量的相關性矩陣Tab. 3 Correlation matrix of monomer amino acid relative abundance in THAA (mol%) in seawater of the low-latitude Northwest Pacific
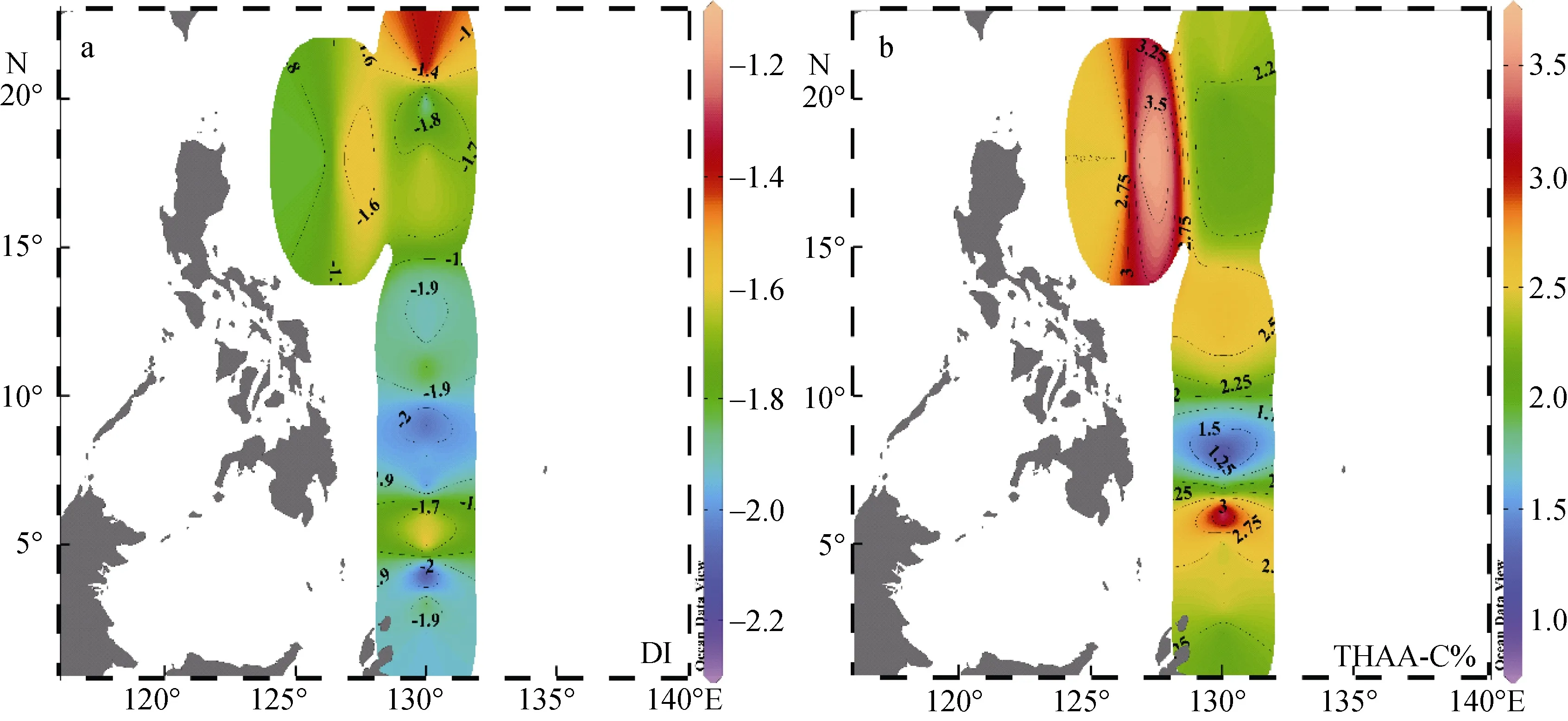
圖7 西北太平洋低緯度海域中DI值 (a)和THAA-C% (b)的水平分布Fig. 7 Horizontal distributions of DI (a) and THAA-C% (b) of the low-latitude Northwest Pacific
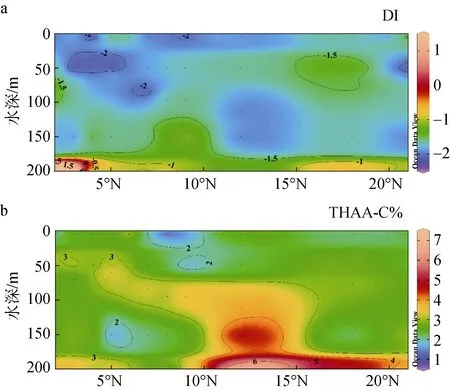
圖8 西北太平洋低緯度海域130°E斷面上層海洋中THAAC%(a)、DI(b)值的垂直分布Fig. 8 Vertical distribution of THAA-C% (a) and DI (b) in the upper ocean (5~200 m) on the 130°E Section of the low-latitude Northwest Pacific
3 結論
西北太平洋低緯度區域表層海水中THAA的濃度范圍為 0.40~0.97 μmol/L, 平均值為 0.58 ± 0.14 μmol/L。THAA整體分布較均勻, 濃度顯著低于中國近海。在近陸NEC區存在高值區, 陸源輸入、以海流為代表的水團運動、浮游植物現場生產等是此海域 THAA的來源, 光降解、細菌降解是此海域 THAA的去除途徑。西北太平洋低緯度區域 5 ~ 200 m 海水中THAA的濃度范圍是0.30 ~ 1.05 μmol/L, 平均值是0.59 ± 0.16 μmol/L。細菌降解作用隨深度增加而減弱和顆粒物沉降過程中的再溶解是使得該海域上層海洋(5 ~ 200 m)中THAA濃度隨深度增加而增加的原因, 而降解因子DI值的變化規律也證明了這一點。對該海域的 THAA組成進行分析后得出, 優勢氨基酸為 Asp、Glu、Ser、Gly、Thr和 Ala, 共占 THAA的 87.74%, 優勢氨基酸種類與該海域的優勢藻類—硅藻有關。THAA與DOC、Chl a等環境參數之間均不存在顯著相關性, 這可能是因為 THAA受到了生物和環流的多重影響, 各環境因子對 THAA施加的影響可能是多因素協同作用, 而具體的影響機制尚需進一步研究。計算得出的兩種降解因子THAA-C%和DI表明, 該海域的DOM生物可利用性遠低于近岸海域, 略高于伊勢灣外的西北太平洋、楚科奇海等海域, 證明 DOM 已遭受較大強度的降解, 且在 5 ~200 m 的深度范圍內, 隨深度的增加, 有機質相對更“新鮮”。
猜你喜歡
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科學(2020年5期)2020-11-26 08:19:22
軟件(2020年3期)2020-04-20 01:45:18
商周刊(2018年15期)2018-07-27 01:41:20
敦煌學輯刊(2018年1期)2018-07-09 05:46:42
北京教育·普教版(2017年1期)2017-02-05 13:26:23
新疆農墾科技(2016年2期)2016-08-21 13:50:16
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
新疆財經大學學報(2015年3期)2015-12-10 03:49:15
