利用野生甘藍改良油菜Ogu CMS恢復材料的菌核病抗性
2020-06-03 09:41:22萬華方丁一娟陳致富梅家琴錢偉
中國農業科學 2020年10期
萬華方,丁一娟,陳致富,梅家琴,錢偉
(西南大學農學與生物科技學院/西南大學農業科學研究院,重慶 400716)
0 引言
【研究意義】甘藍型油菜(Brassica napusL.,AACC,2n=38)是由白菜(Brassica rapaL., AA,2n=20)和甘藍(Brassica oleraceaL.,CC,2n=18)雜交,并經自然加倍演化而來,是世界上重要的糧油作物和能源作物。中國是世界第二大油菜種植國,其長江流域油菜產業常年受到腐生真菌核盤菌(Sclerotinia sclerotiorum(Lib.) de Bary)的嚴重危害。由于現有油菜中缺乏菌核病的有效抗源,抗病甘藍的發現為油菜抗病育種帶來了新的思路。細胞質雄性不育(cytoplasmic male sterility,CMS),尤其是蘿卜胞質不育系Ogu CMS的不育性十分穩定,是蕓薹屬作物的安全授粉控制系統[1],也是生產油菜雜交種最好利用的授粉控制系統之一。因此,以含有野生甘藍優良抗病遺傳成分的人工合成甘藍型油菜為橋梁,培育抗菌核病的油菜Ogu CMS恢復系具有重要理論意義和實踐意義。【前人研究進展】由于油菜基因庫中匱乏高抗或免疫的資源[2-5],學者們已嘗試從油菜的近緣物種中去發掘優良抗源[5-7]。作為油菜的親本物種之一,甘藍遺傳多樣性豐富[8-10],并且由于其長期受到野生環境的自然選擇,存在優良的抗病資源。野生甘藍B.incana具有良好的菌核病抗性,其莖稈抗病性可達到耐病油菜對照中油821的20倍[7],其抗病QTL(quantitative trait locus)分布在C1、C3、C4、C7和C9等多條連鎖群[11],與現已報道的油菜抗菌核病QTL位置不同[12-14],其中,部分與甘藍抗病QTL連鎖的標記已成功應用于抗菌核病分子標記輔助選擇[15-16]。為將此野生甘藍的菌核病抗病位點導入油菜,DING等[17]以其為抗源,與白菜進行遠緣雜交,發展了人工合成甘藍型油菜,表現出較好的菌核病抗性。【本研究切入點】油菜中已成功育成了甘藍型油菜Ogu胞質不育系[18-19]及Ogu CMS的恢復系[20-22],但受限于油菜抗源的匱乏,目前還沒有抗菌核病的油菜 Ogu CMS恢復系報道。【擬解決的關鍵問題】本研究以人工合成甘藍型油菜橋梁,將野生甘藍的抗菌核病位點轉入油菜,發展具有甘藍抗病遺傳成分且含恢復基因的材料,為發展抗菌核病油菜Ogu CMS恢復系提供種質資源。
1 材料與方法
1.1 試驗時間和地點
田間試驗于 2013—2016年在重慶市油菜工程技術研究中心試驗基地和內蒙古大學試驗基地(夏季加代繁殖)進行。
1.2 試驗材料及選育策略
前期以高抗菌核病野生甘藍B. incana(編號C01)與白菜(編號6Y733)雜交,經胚挽救、染色體加倍獲得了抗菌核病的人工合成甘藍型油菜AC1-4(相對于中雙9號的枝稈和葉片,相對感病度S分別為0.75和0.57,雙高品質)。以AC1-4為父本,對油菜Ogu CMS胞質不育雜種4Q292(源于Ogu CMS油菜與常規油菜的雜種F1,感菌核病,雙低品質)人工授粉,通過對后代單株進行抗病位點連鎖標記輔助選擇(marker-assisted selection,MAS)、可育株抗性鑒定、結實率統計和品質分析,從中選擇可育株,自交發展F2代;對F2代單株根據上述方式篩選攜帶野生甘藍抗病QTL、且其他性狀較優的個體自交;在自交后代中繼續重復上述篩選,最后獲得攜帶野生甘藍抗病QTL、育性正常、產量和品質相對較優的 Ogu CMS恢復系材料。
1.3 分子標記輔助選擇(MAS)
以野生甘藍 C01的帶型為參考,利用 4Q292和AC1-4對前期研究中鑒定到的野生甘藍所有抗病QTL區間及側翼最近SSR標記進行篩選,獲得7個在雙親之間存在多態性的標記,涉及3個葉片抗性位點和2個莖稈抗位點(表 1)。利用表中標記對各世代單株進行分析。盡管其抗病QTL分布在C1、C3、C4、C7和C9等多條連鎖群[11],但C1和C9聯鎖群上的2個QTL(C1-QTL和 C9-QTL)解釋的表型變異度相對較高(13.6%—24.1%),為主效QTL,且QTL均具有至少2個連鎖的多態性標記,因此,MAS過程中首要選擇同時攜帶C1和C9抗病位點,其次考慮是否攜帶C3和C7上QTL連鎖標記(對表型貢獻率相對較低,且均為單標記選擇)。MAS于苗期在每個世代開展,經MAS選中的單株進入后期的抗性鑒定、結實性調查和品質分析。
1.4 菌核病抗性鑒定
核盤菌菌種為 1980。參照 MEI等[23]的方法進行菌種活化、接種體(PDA菌絲塊)制備及菌核病抗性鑒定。于油菜終花期后2周進行離體枝稈菌核病抗性鑒定:取一次有效分枝,截取長約30 cm一段,將兩端用保鮮膜纏繞(以防止水分散失),然后轉移到可控溫、控濕的室內,將枝稈隨機擺放于鋪好濕毛巾的接種臺上,并將準備好的PDA菌絲塊帶菌面緊貼于枝稈上,每枝稈接種2—3個菌絲塊。接種完,在接種臺上放上支架,以塑料膜密封,控制溫度((22±1)℃)和濕度(90%—95%)。接種后第 4天測量菌斑的長度,以相對于耐病對照甘藍型油菜中雙9號的感病度(S)評價材料菌核病抗性,即S=L待測/L對照(L待測表示材料接種核盤菌的病斑長度(cm),L對照表示耐病對照中雙9號的病斑長度(cm))。
1.5 結實率統計及籽粒品質分析
每個單株分別選取主花序上連續 50個自交角果和離主花序最近且未套袋一次分枝上 50個自由授粉角果,統計每角果籽粒數,分別代表自交和自由授粉結實率。用FOSS NIR system3700型近紅外光譜儀檢測自交種籽粒品質。光譜掃描條件:分辨率8 cm-1、掃描次數64次、光譜范圍1 100—2 500 nm、溫度20—25℃。
1.6 統計分析
采用SAS 8.0軟件進行數據分析。
2 結果
以高抗菌核病的野生甘藍C01為父本,與編號為6Y733的白菜雜交,10 d后取子房解剖后進行胚挽救及染色體加倍,獲得人工合成甘藍型油菜AC1-4,其苗期形態介于野生甘藍(圖 1-A)和白菜(圖 1-B)之間,與甘藍型油菜形態相似(圖 1-C),分子標記也表明,AC1-4為B. incana和6Y733的真雜種(圖2)。對雜種F1進行體細胞染色體觀察發現,單倍體植株均含19條染色體(圖1-F),經染色體加倍的AC1-4植株體細胞均含有38條染色體(圖1-G)。花粉育性鑒定結果表明,單倍體植株的花粉育性(圖 1-H)遠低于人工合成甘藍型油菜AC1-4的花粉育性(可達80%)(圖1-I)。葉片和枝稈菌核病抗性鑒定結果(圖1-J和圖1-K)表明,人工合成甘藍型油菜AC1-4的葉片抗性和枝稈抗性都顯著高于對照甘藍型油菜中雙9號和白菜親本。

圖1 高抗菌核病人工合成甘藍型油菜AC1-4的特征Fig. 1 Characteristics of AC1-4 (resynthesized Brassica napus) with high resistance against Sclerotinia sclerotiorum

圖2 SSR引物SWU260檢測親本及F1Fig. 2 Verification of the relatives and F1 hybrid individual plants with SSR primer SWU260
人工合成甘藍型油菜AC1-4與Ogu CMS甘藍型油菜雜種4Q292(圖1-D)雜交產生的F1代(圖1-E)代共獲得146個單株,其中69株可育,77株不育。經卡方檢驗,F1代育性符合1∶1分離,(χ21∶1= 0.17,P>0.05),符合預期分離比。對F1代146個單株進行分子標記輔助選擇,其中128個單株遺傳了源于抗病甘藍B. incana的C1-QTL和C9-QTL,另外分布在C3和C7染色體的2個標記也在上述128個單株中表現出AC1-4與4Q292的雜合帶型。剩余的18個單株中,有 11個單株丟失了 C1-QTL,2個單株丟失了C9-QTL,5個單株的C3或C7-QTL連鎖標記未表現出抗病帶。F1代植株的自交結實率與天然結實率顯著相關(r= 0.52),但套袋自交的角果粒數變異較大,最多達24粒/角果,最少為5粒/角果,平均為9.07粒/角果。菌核病抗性鑒定表明,含有所有標記抗病帶的49個F1可育單株的相對感病度S介于0.2—1.4,平均值為0.74,而不完全遺傳上述7個標記抗病帶的6株可育株相對感病度S介于0.9—1.5,平均值為1.23。結合結實率、分子標記結果以及收獲種子狀況(種子量及飽滿度),選取編號為4C99和4C115的2個F1單株進入下一輪篩選,2份材料的相對感病度S分別為0.66和0.40,自交結實率分別為12.4和15.0粒/角果,自由授粉結實率分別為17.9和21.4粒/角果。
將4C99和4C115的自交種進行了夏繁種植,但由于環境條件的變化,導致大量材料F2單株未開花,最終4C99自交后代有59株開花,4C115有65株開花。對上述124個F2單株進行C9上枝稈主效抗病QTL連鎖標記篩選,發現源于4C99的59個F2單株中有35個植株攜帶 C9-QTL,源于4C115的F2單株中有34個植株攜帶C9-QTL。由于夏繁地無抗性鑒定條件,因此,未對F2代植株進行抗性鑒定和篩選。隨后對可育株上獲得自交種的材料進行了籽粒品質分析,最終結合標記分析及品質分析數據篩選了8個F2,其芥酸含量介于7.50%—24.70%,硫苷含量介于15.88—78.63 μmol·g-1餅粕。利用該8個F2單株進行自交發展下一代,田間種植共獲得的8個F2:3家系(以5X進行編號),家系大小介于46—184株。利用分子標記對8個家系進行分析(表2),F2:3代材料基因型分離明顯,無論是C1還是C9上的QTL在各個家系中均存在分離,其中4個家系(5X78、5X80、5X82和5X84)中出現同時攜帶C1-和C9-QTL的概率(36.6%— 43.5%)顯著高于其他4個家系(0—8.7%)。8個家系共計獲得222株同時攜帶C1-和C9-QTL的植株。為了進一步縮小選擇范圍,利用所有抗病標記結果進行了各家系材料與抗病親本的聚類分析,最終從5X78、5X80、5X81、5X82、5X83和 5X84中分別篩選到 4株、23株、2株、16株、1株和4株(共計50株)與抗性親本遺傳關系較近的可育株,而5X77和5X79 2個家系中沒有發現與抗性親本聚在一起的植株。對此50份材料進行枝稈菌核病抗性鑒定,同時以25株完全或部分缺失了抗病QTL的材料作為對照,結果顯示,50份材料的相對感病度S介于0.34—0.89,平均值為0.60,而完全或部分缺失了抗病QTL的植株平均相對感病度S為0.98,顯著高于前者(P<0.01)(圖3)。對部分材料進行了籽粒品質檢測(表3),發現所有材料均表現為雙高品質。盡管如此,材料之間的硫苷和芥酸含量表現出分離狀態,個別材料如5X82-2和 5X82-7的菜籽品質已較為接近單低水平,其中5X82-2 的硫苷含量為 45.38 μmol·g-1餅粕,5X82-7 的芥酸含量為6.5%。同時,這兩個材料的相對感病度S分別為0.33和0.48(表3),表明其具有單低的品質,且菌核病抗性已得到改良。結合 MAS和枝稈抗性鑒定結果,上述材料中相對感病度S不超過0.5的單株自交種將進入下一輪篩選。
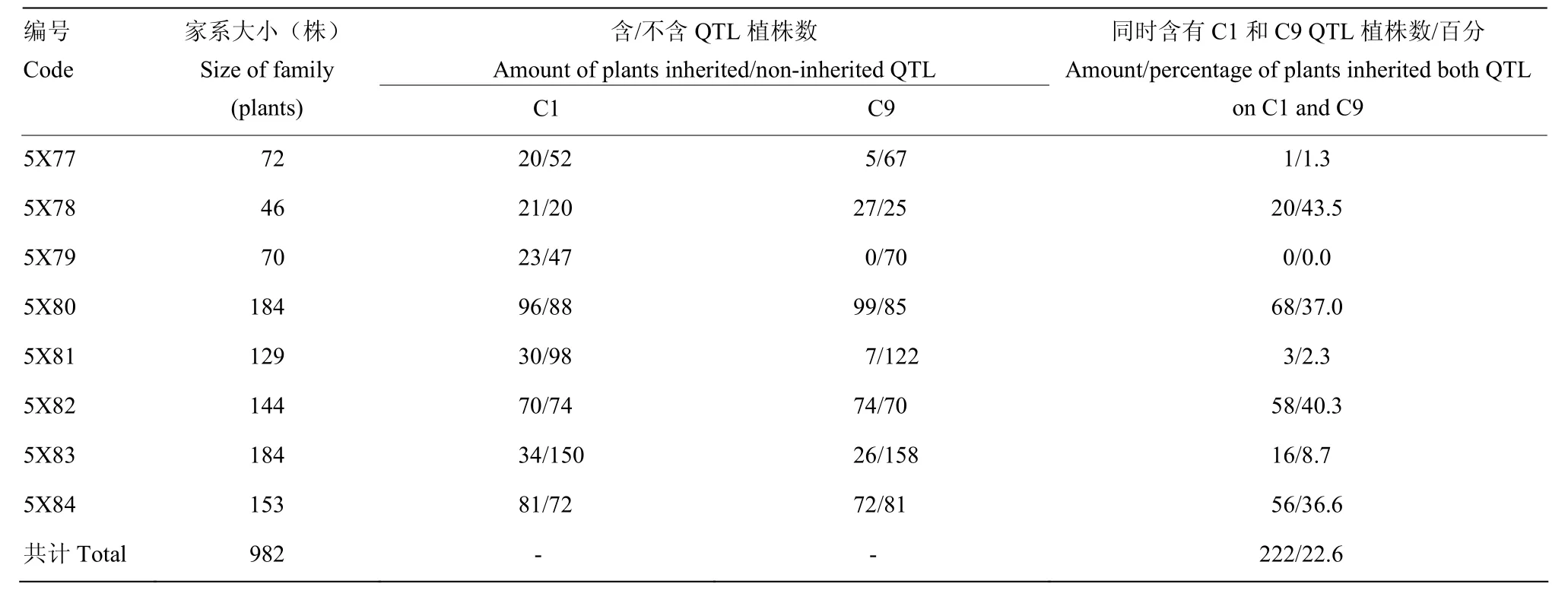
表2 各F2:3家系分子標記分析結果Table 2 Results from molecular marker analysis of F2:3 families
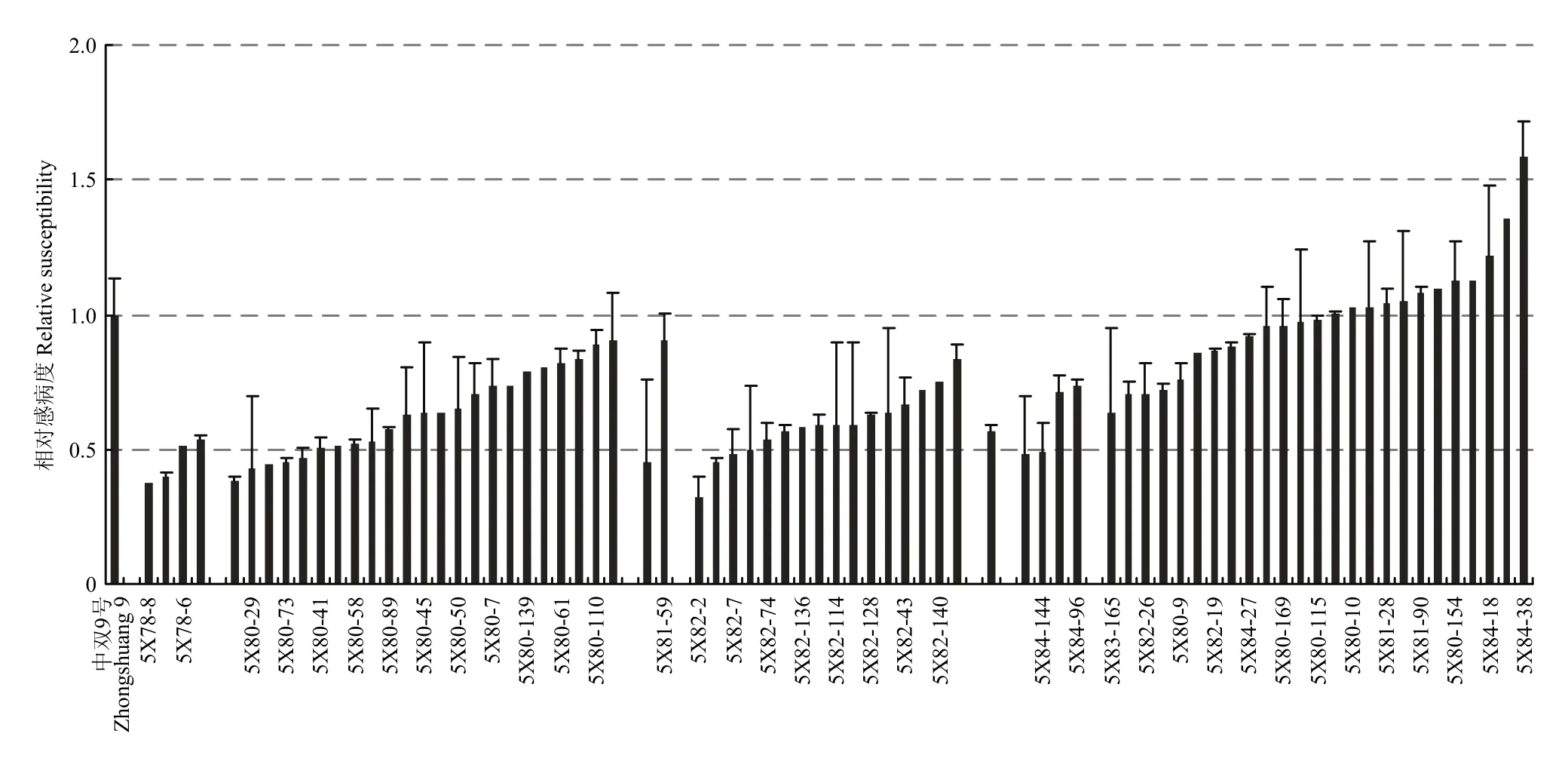
圖3 各F2:3家系中選植株的菌核病抗性Fig. 3 The Sclerotinia resistance level of selected individuals in F2:3 families
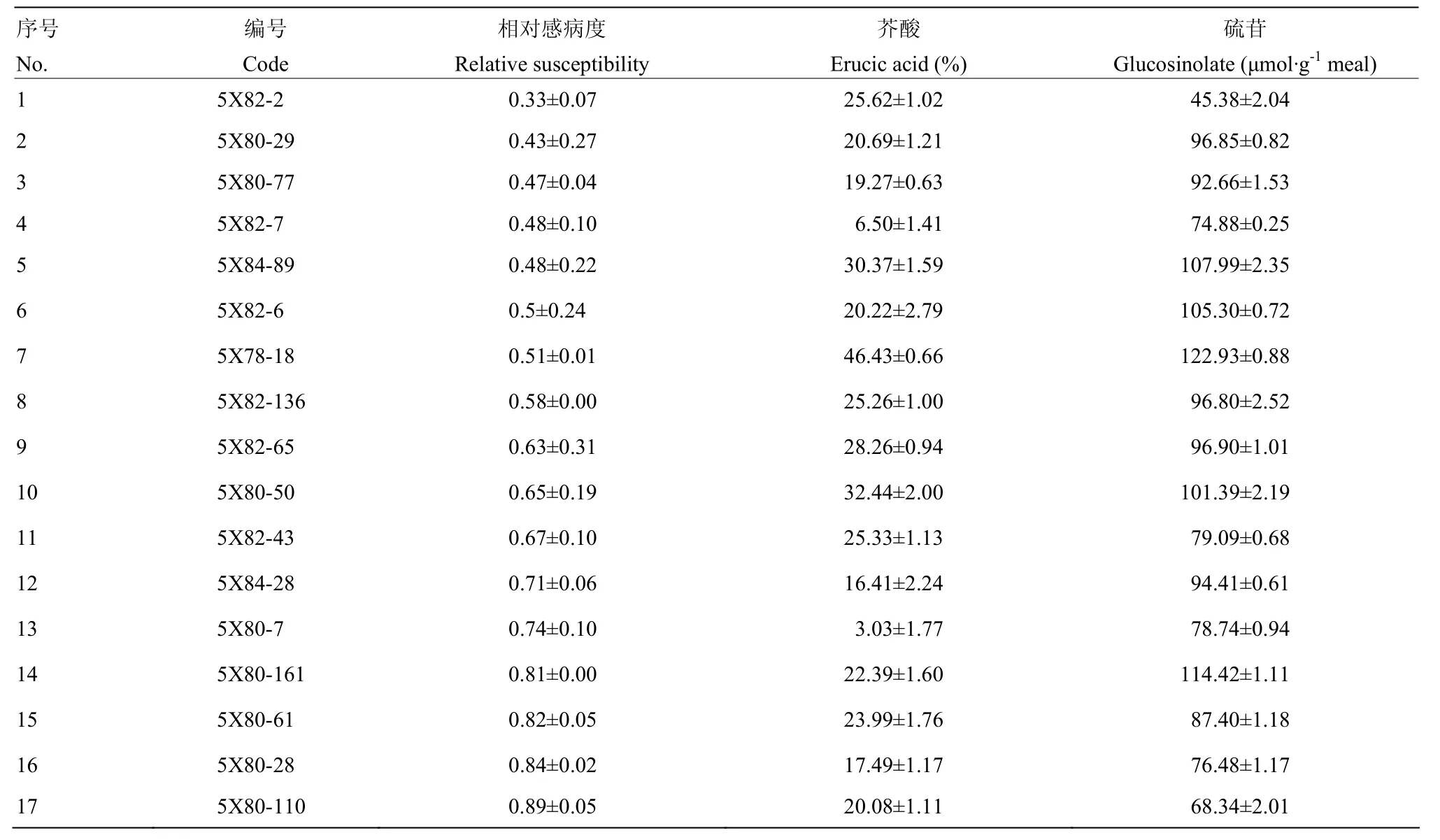
表3 F2:3家系中部分單株抗性和籽粒品質Table 3 Relative susceptibility and seed quality of some individuals in F2:3 generation
3 討論
甘藍型油菜的遺傳背景因“雙低”選育過程而趨于狹窄,但其二倍體祖先種白菜和甘藍起源早、栽培歷史悠久,形態、農藝性狀具有豐富的變異[24]。利用甘藍和白菜培育的人工合成甘藍型油菜,已被廣泛運用到甘藍型油菜遺傳育種,比如胞質雄性不育系“Bro CMS”創制[25]、大粒甘藍型油菜創制[26],黃籽甘藍型油菜品系創制[27-28]、早熟性狀改良[29]、黃萎病抗性的種質創建[30]、根腫病抗性改良[31]以及Ogu育性恢復基因(Rfo)轉育[32]等研究中。人工合成甘藍型油菜與天然甘藍型油菜遺傳差異大,可有效拓寬天然甘藍型油菜的遺傳資源。然而,人工合成甘藍型油菜的含油量較低,芥酸、硫苷含量較高,甚至出現一些源于祖先種的不良性狀,常需要不斷進行品質改良。本研究所使用的 AC1-4也表現出雙高品質,以其為親本發展的低世代中出現芥酸或硫苷含量較高的材料。后續研究中,將進一步發展高世代,進行雙低品質材料的選擇。
Ogura細胞質雄性不育敗育徹底、轉育容易,是蕓薹屬作物的安全授粉控制系統[1]。甘藍型油菜 Ogu胞質不育系[18-19]及 Ogu CMS的恢復系[20-22]已成功創建,但目前油菜基因庫中缺乏有效抗源,限制了抗菌核病的油菜Ogu CMS恢復系的研究。甘藍型油菜的祖先種甘藍是改良油菜菌核病抗性的優良資源[11,33]。在早期研究中,鑒定到野生甘藍B. incana具有較好的菌核病抗性[7],并對其抗病位點進行了QTL定位[11]。利用該材料我們創制了人工合成甘藍型油菜,該人工合成材料理論上應該遺傳B. incana的整套C基因組染色體,因此,該人工合成系與Ogu CMS油菜的雜種F1所有植株理應繼承B. incana的抗病QTL位點。然而,利用QTL區間或側翼標記對F1代的標記分析后發現,有極少數單株存在個別QTL丟失的情況。這可能是由人工合成甘藍型油菜的遺傳不穩定造成的:人工合成甘藍型油菜含有A、C兩套共線性較高的基因組,減數分裂時可能產生A、C染色體之間的異源配對[34-35],形成多價體及單價體[36-37],從而導致染色體分離紊亂和非整倍體配子的產生[38-39],使某些遺傳信息在后代可能丟失。
本研究利用人工合成系和Ogu CMS油菜尋找到7個與抗病QTL連鎖的多態性標記,涉及4個染色體上5個抗病QTL(包括葉片抗性QTL和莖稈抗病QTL)。因此本研究首先采用C1和C9抗病QTL兩側均有多態性的標記對材料進行分析和選擇,而其他3個QTL只開放出一個單標記,這些標記僅用于輔助選擇和遺傳分析,而用于MAS的準確性低。油菜及甘藍的菌核病抗性均為數量性狀,主要受到加性效應控制[11-14,40],因此,本研究在每個世代盡量篩選同時具有C1和C9 QTL、且盡量在其余3個標記結果中選擇具有抗病甘藍帶型的個體。后期的抗性鑒定顯示,標記選中的材料平均抗性顯著高于 QTL出現丟失的材料,一定程度證明了標記選擇的有效性。
結合 MAS和抗性選擇,盡管在每個世代均能鑒定到同時攜帶多個抗病QTL且相對感病度S較低的單株,但是從對 F2:3家系的分析中可以看出,所有家系中的單個抗病位點均未達到純合,且硫苷和芥酸含量也表現出分離。但是,將根據 1.2的選育策略對 F2:3中選擇到的候選植株進行進一步的選擇,且后期的選擇將注重以家(株)系為單位進行分析,以確定目標位點及控制育性的核基因Rf是否純合;同時需要增加家(株)系中個體數,從而出現更多的交換重組,從中獲得既具有抗病位點、又具有低硫苷和低芥酸位點的重組個體;再者,可以考慮通過目前獲得的抗病且芥酸或硫苷含量接近單低的個體相互雜交,聚合不同性狀的優良位點,從而獲得優質且抗性提高的恢復株系。
4 結論
以人工合成甘藍型油菜為橋梁,野生甘藍B.incana的菌核病抗性可被有效轉移到甘藍型油菜,從而改良油菜Ogu CMS恢復材料的菌核病抗性。
