雷帕霉素靶蛋白復合物1對腸道3型固有淋巴細胞功能的影響
2020-05-28 05:47:56劉健悅
上海交通大學學報(醫學版) 2020年4期
劉健悅 ,沈 蕾
1.上海交通大學基礎醫學院免疫學與微生物學系,上海200025;2.上海市免疫學研究所,上海200025
3型固有淋巴細胞(group 3 innate lymphoid cell,ILC3)是近年來新發現的一類重要的免疫細胞亞群,主要富集在腸道黏膜組織,表達轉錄因子視黃酸受體相關的孤兒核受體(retinoic acid receptor related orphan receptor,RORγt),并分泌細胞因子白細胞介素22(interleukin-22,IL-22)和IL-17[1-3]。ILC3表面缺乏抗原特異性受體的表達,在腸道中主要受細胞因子IL-23/IL-1β調控激活,在維持黏膜屏障完整性和抗病原體感染過程中發揮著重要作用[4-5]。研究[6]表明,ILC3功能缺失或低下可導致腸道細菌感染。在小鼠中,ILC3可基于細胞表面標志物趨化因子受體6(C-C motif chemokine receptor 6,CCR6)和天然細胞毒性受體(natural cytotoxicity triggering receptor,NCR,又稱NKp46)的差異表達劃分為不同亞群[7]。其中,CCR6+NKp46-ILC3是腸道淋巴組織發育所需的,可分泌IL-22和IL-17A;而共表達RORγt和T-bet(T-box transcription factor 21)的CCR6-NKp46+ILC3和CCR6-NKp46-ILC3主要分泌IL-22[8-9]。IL-22對于組織修復和宿主腸道組織抗細菌感染至關重要。一方面,IL-22信號可促進上皮細胞增殖、細胞間緊密連接的形成以及傷口愈合;另一方面,IL-22還調控與抗菌肽形成有關的基因表達以及上皮細胞的巖藻糖基化[10-13]。近年來,對ILC3發育和功能的研究越來越多,揭示了其在機體免疫中的重要性;但作為新興領域,仍有許多問題需要解決,關于ILC3穩態及功能調節的相關研究仍具有重要意義。
雷帕霉素靶蛋白(mechanistic target of rapamycin kinase,mTOR)是一種進化上高度保守的非典型絲氨酸/蘇氨酸蛋白激酶,屬于磷酸肌醇-3-激酶(phosphatidylinositide 3-kinase,PI3K)相關激酶家族,主要以2種復合體mTORC1(mTOR complex 1)和mTORC2的形式存在[14-15]。其中,關于mTORC1活化和功能的研究更為廣泛和透徹,其可整合生長因子、壓力、能量狀態及氧氣等多種胞內外信號,調控細胞生長、增殖、分化和自噬等過程。mTORC1主要由Raptor(regulatory associated protein of mTOR)、mLST8(mTOR associated protein)、PRAS40(proline-rich Akt substrate 40 kDa)、DEPTOR(DEP domain containing mTOR interacting protein)以及mTOR分子組成,對雷帕霉素反應敏感[16-18]。越來越多的研究表明mTORC1對免疫細胞的發育和功能起著重要的調控作用。mTORC1可以通過影響轉錄因子STAT(signal transducer and activator of transcription)家族成員從而決定CD4+T細胞的譜系分化,如Yang等[19]發現mTORC1的重要組分Raptor缺失后會導致Th2和Th17細胞分化受損,但不影響Th1細胞的分化。mTORC1還可通過作用于缺氧誘導因子1(hypoxia inducible factor 1,HIF1)和干擾素調節因子4(interferon regulatory factor 4,IRF4)促進CD8+效應T細胞的分化和功能[20]。在B細胞中,有研究[21]發現mTORC1功能被抑制后,B細胞增殖和漿細胞分化減弱,體液免疫應答受損。此外,mTORC1還參與調節IL-23誘導的中性粒細胞中IL-22和IL-17的產生[22]。然而目前為止,關于mTORC1對固有淋巴細胞調控的研究仍很少。鑒于固有淋巴細胞與T細胞之間的相似性,我們推測mTORC1在ILC3的發育維持和功能調控中也起著重要作用。
本研究先通過體外的雷帕霉素(rapamycin)抑制實驗探究mTORC1對ILC3功能的調控作用,然后構建ILC3特異性mTORC1功能缺失的條件性敲除小鼠,檢測該小鼠腸道ILC3發育及功能的變化情況。
1 材料與方法
1.1 實驗材料
1.1.1 實驗動物 本研究中所有小鼠均為C57BL/6背景,Rptorflox/floxRorcCre(Rptor為編碼Raptor蛋白的基因,Rorc為編碼RORγt蛋白的基因)小鼠由Rptorflox/flox小鼠和Rorccre轉基因小鼠雜交獲得,其中Rptorflox/flox小鼠受贈于中國人民解放軍第三軍醫大學葉麗林教授課題組,Rorc-cre轉基因小鼠受贈于中國科學院上海營養與健康研究所邱菊教授實驗室。野生型C57BL/6小鼠購自上海斯萊克實驗動物有限公司。所有實驗小鼠均飼養于上海交通大學醫學院醫學實驗動物科學部的SPF級設施中,實驗動物使用許可證為SYXK(滬)2018-0027。除非本文另有說明,否則實驗用小鼠均為同窩6~8周齡雌鼠。所有動物實驗均經上海交通大學醫學院動物保護委員會批準。
1.1.2 主要試劑與儀器抗CD3、CD45R(B220)、CD11b、CD11c、Ly-6G(lymphocyte antigen 6 complex,locus G)、CD45.2和IL-22的抗體均購自BioLegend公司(美國),抗RORγt、NK1.1(CD161)和CD90.2抗體購自eBioscience公司(美國),抗殺傷細胞凝集素樣受體G1(killer cell lectin like receptor G1,KLRG1)抗體購自BD Biosciences公司(美國),蛋白轉運抑制劑Brefeldin A和細胞破膜固定液購自Thermo Fisher公司(美國),DMEM培養基和胎牛血清購自Gibco公司(美國),膠原酶Ⅷ和脫氧核糖核酸酶Ⅰ(DNaseⅠ)購自Sigma Aldrich公司(美國)。另有試劑IL-23(R&D Systems,美國)、雷帕霉素(Selleck Chemicals,美國)、小鼠調節性T細胞染色試劑盒(eBioscience,美國)、PrimeScript RT試劑盒(TaKaRa,日本)、SYBR Green核酸熒光染料(Applied Biosystems,美國)。實驗中所用到的儀器主要有BD FACSAriaTMⅢ流式分選儀(BD Biosciences,美國)、BD LSRFortessaTMX-20流式分析儀(BD Biosciences,美國 )、7500 Fast Real-time PCR儀(Applied Biosystems,美國)。
1.2 實驗方法
1.2.1 小鼠腸道固有層淋巴細胞的分離 小鼠脫頸處死后取出腸道,剝除上面附著的脂肪,剪去派氏結(Peyer′s patch),沿縱向剪開,放入磷酸鹽緩沖液(PBS)中震蕩清洗,去除糞便。之后剪成1~2 cm的小段,先放入含有1 mmol/L二硫蘇糖醇(DTT)的PBS中震蕩10 min,再轉移到含30 mmol/L乙二胺四乙酸(EDTA)和10 mmol/L 4-羥乙基哌嗪乙磺酸(HEPES)的10 mL PBS中震蕩10 min。然后用含有0.05% DNaseⅠ和100 U/mL膠原酶Ⅷ的4 mL RPMI 1640培養基37 ℃培養90 min。消化結束后劇烈晃動使組織細胞分散并用孔徑100 μm的細胞過濾器過濾,然后在室溫下以500×g離心20 min,從80%和40%Percoll梯度液的中間層收獲單個核細胞,得到腸道固有層淋巴細胞(lamina propria leukocyte,LPL)。
1.2.2 流式染色 得到的腸道LPL先按1:100加入Fc端封閉劑(CD16/32),4 ℃孵育10 min。細胞表面蛋白如CD3、CCR6、CD11b、CD45、CD11c等的染色按1:200比例稀釋抗體,4 ℃避光孵育30 min,然后PBS洗去抗體。對于細胞內部蛋白如IL-22及核內轉錄因子如RORγt的染色,先用小鼠調節性T細胞染色試劑盒將細胞固定透化1 h,再按1:100比例稀釋胞內抗體,4 ℃避光孵育1 h,PBS洗去抗體,最后用200 μL PBS重懸細胞。過程中離心程序均為400×g,5 min。
1.2.3 實時熒光定量PCR 分選獲得的細胞先用TRIzol試劑抽提出RNA,檢測RNA濃度,并用PrimeScript RT試劑盒反轉錄成cDNA。使用SYBR Green試劑盒和不同的引物組進行實時熒光定量PCR(real-time qPCR),每個樣本設置3個復孔,檢測程序為95 ℃ 10 min;95 ℃ 15 s,60 ℃1 min,95 ℃ 15 s,40個循環;60 ℃ 1 min。結果是相對于次黃嘌呤磷酸核糖轉移酶(hypoxanthine phosphoribosyl transferase,Hprt)基因標準化的相對表達值,并根據2-ΔΔCT方法統計。實驗中所用到的引物序列見表1。
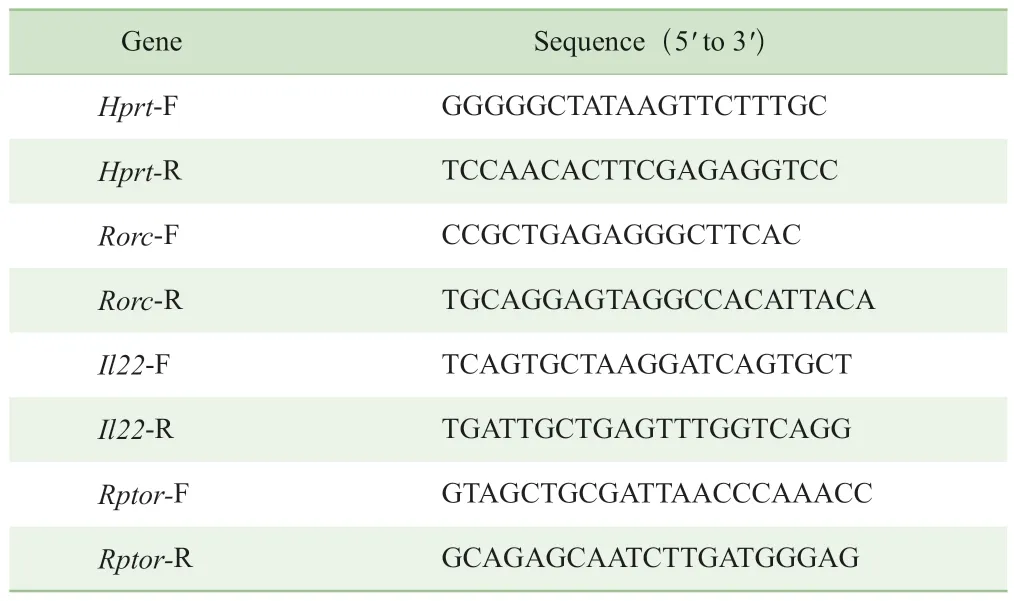
表1 Real-time qPCR引物Tab 1 Primer sequences for real-time qPCR
1.3 統計學分析
本實驗中的數據使用GraphPad Prism 7.0軟件進行統計學分析,定量資料用±s表示;2組間差異的比較采用非配對Student′st檢驗計算。統計圖中所用樣本數據均來源于2個及以上獨立的實驗結果,P<0.05時認為差異具有統計學意義。
2 結果
2.1 體外雷帕霉素刺激抑制ILC3分泌IL-22
雷帕霉素是mTORC1信號通路的特異性抑制劑,對野生型C57BL/6小鼠腸道LPL用雷帕霉素體外刺激6 h,流式細胞分析檢測ILC3數量(RORγt+細胞)及IL-22的分泌情況。結果顯示0.1 mg/mL雷帕霉素處理后,ILC3比例及數量無變化,但分泌的IL-22明顯減少(圖1);圖中也可觀察到RORγt陰性的非ILC3細胞IL-22產生能力不受影響,這提示mTORC1對ILC3功能具有調控作用。

圖1 雷帕霉素對ILC3分泌IL-22的抑制作用Fig 1 Inhibitory effect of rapamycin on IL-22 secretion of ILC3
2.2 IL-23刺激后ILC3中Rptor mRNA水平表達上調
上述rapamycin刺激LPL實驗中,無法排除LPL中其他淋巴細胞對ILC3的干擾作用。故取6~8周齡野生型小鼠腸道LPL,借助流式分選技術獲取純化的ILC3。小鼠腸道ILC3的分選策略為:首先借助側向散射光(side scatter,SSC)去除成團的細胞,然后根據前向散射光(forward scatter,FSC)和SSC確定淋巴細胞亞群的位置,再利用譜系標記(lineage marker)(CD3、B220、CD11b、CD11c、Ly-6G)定義出Lin-淋巴細胞群,之后通過CD45.2和CD90.2圈出CD45.2lowCD90.2high細胞群,ILC3主要集中在這一群中,最后使用KLRG1和NK1.1排除其中少量的ILC2及ILC1,得到純化的ILC3(圖2A)。對分選獲得的ILC3進行RORγt表達水平的回測,結果證實分選得到的ILC3中RORγt陽性的比例達到90%以上(圖2B)。
IL-23是激活ILC3的主要上游信號,活化后的ILC3可分泌更多的IL-22等細胞因子,調節腸道穩態。將利用上述方法分選出來的ILC3用10 ng/mL IL-23刺激24 h,然后通過real-time qPCR檢測Rorc和Il22的mRNA表達水平,發現與已有的研究結果一致,這2種基因的mRNA表達均上調。結果還顯示,mTORC1的關鍵組成蛋白Raptor的mRNA表達水平也同步上調(圖2C)。上述結果提示,mTORC1可能在ILC3分泌IL-22的過程中起調控作用。
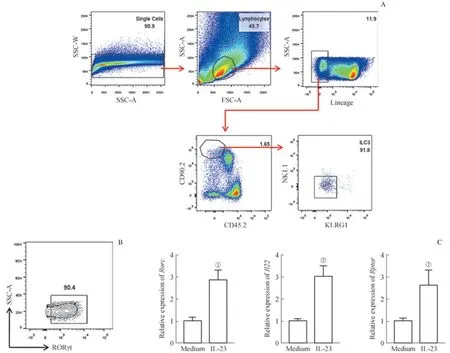
圖2 ILC3的純化及IL-23對其Rorc、Il22和Rptor mRNA表達的影響Fig 2 Purification of ILC3 and effect of IL-23 on Rorc, Il22 and Rptor mRNA expression
2.3 mTORC1功能缺失不影響小鼠腸道ILC3的數量
為了進一步探究mTORC1在小鼠腸道ILC3中的作用,用Rptorflox/flox小鼠(下文簡稱為WT小鼠)與Rorc-cre轉基因小鼠雜交,從而獲得只在表達RORγt的細胞中特異性敲除Rptor基因的小鼠Rptorflox/floxRorcCre(下文簡稱為KO小鼠),該小鼠是研究mTORC1功能缺失常用的工具小鼠。首先用real-time qPCR驗證Rptor的敲除效率,KO小鼠ILC3中RptormRNA表達水平顯著降低,說明條件性敲除小鼠構建成功(圖3A)。
借助流式細胞技術進一步分析小鼠結腸ILC3及ILC3中各亞群的比例及數量,發現WT小鼠和KO小鼠之間并無明顯差異(圖3B、C)。隨后,又比較了穩態下WT小鼠和KO小鼠小腸及結腸長度和腸道派氏結的數量,仍無明顯差異(圖3D、E)。最后,通過蘇木精-伊紅染色法(hematoxylin-eosin staining,H-E染色)對比了WT小鼠和KO小鼠結腸結構,結果顯示特異性敲除Rptor后并未使穩態下腸道結構發生明顯改變(圖3F)。

圖3 KO小鼠與WT小鼠ILC3及ILC3中各亞群的比例及數量和腸道結構觀察Fig 3 Ratios and numbers of ILC3 and ILC3 subsets and observation of intestinal structure in KO mice and WT mice
2.4 mTORC1功能缺失抑制ILC3分泌IL-22的能力
鑒于上述KO小鼠結腸的ILC3數量沒有發生變化,后續實驗檢測了KO小鼠結腸ILC3活化后分泌細胞因子的能力是否發生改變。流式細胞分析結果顯示,在使用1 ng/mL IL-23刺激8 h后,相比WT小鼠,KO小鼠分泌IL-22的量顯著減少(圖4A),但分泌粒細胞巨噬細胞集落刺激因子(granulocyte macrophage colony stimulating factor,GMCSF)的能力不變(圖4B)。相應地,在分選出WT小鼠和KO小鼠腸道ILC3后,使用1 ng/mL的IL-23刺激8 h,通過real-time qPCR檢測Il22mRNA及編碼GM-CSF的基因Csf2mRNA水平,結果與流式細胞分析結果一致(圖4C)。
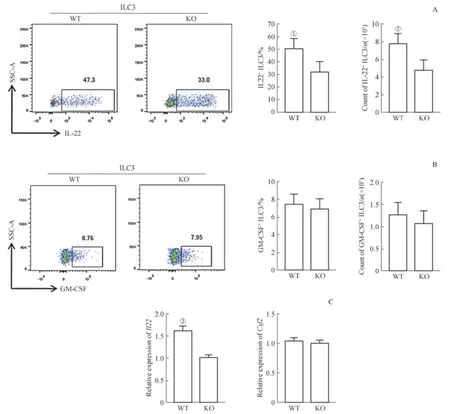
圖4 Raptor缺失后ILC3分泌IL-22的能力Fig 4 Ability of ILC3 to secrete IL-22 after Raptor-deficiency
3 討論
腸道免疫系統穩態的維持依賴于分布在腸黏膜上皮和固有層的各類免疫細胞協同作用。ILC3是近年來發現的一類免疫細胞亞群,主要富集于腸道黏膜組織,在腸道免疫穩態和微生物防御中發揮重要作用[1-2]。在腸道生理穩態下,共生微生物來源的相關信號首先促使巨噬細胞及樹突狀細胞等分泌IL-23和IL-1β,兩者作為ILC3的主要上游信號,可使ILC3活化并分泌細胞因子IL-22和IL-17A等;其中,IL-22被認為對腸道屏障穩態起著正向調控作用。IL-22主要參與促進上皮細胞抗菌肽及黏液的生成、上皮細胞增殖和巖藻糖基化以及組織修復等過程,從而維持腸道屏障穩態[10-12]。研究[8]表明,ILC3中IL-22分泌缺失或低下可導致腸道細菌感染。ILC3和輔助性T細胞(Th17、Th22)均可產生IL22,但ILC3已被證實是腸道穩態下IL-22的主要來源。關于ILC3中IL-22分泌的調控已有報道,主要集中在轉錄水平,如芳香烴受體(aryl hydrocarbon receptor,Ahr)及轉錄因子STAT3等都參與誘導IL-22的生成[23-24],但仍存在很多未知的調控機制。
本研究先借助體外雷帕霉素刺激LPL實驗發現mTORC1對ILC3功能具有調控作用,又在IL-23刺激ILC3活化過程中觀察到編碼IL-22和mTORC1關鍵組分蛋白Raptor的基因mRNA表達水平同步上調,猜測mTORC1可能參與調控ILC3中IL-22的生成。通過構建Rptorflox/floxRorcCre特異性基因敲除小鼠,發現ILC3缺失Raptor后其分泌IL-22的能力受損,但并未影響小鼠結腸ILC3及ILC3亞群的數量,這說明mTORC1調控了ILC3的功能,但并未影響ILC3的發育。同時腸道穩態下在KO小鼠中未觀察到腸道結構損傷,這可能是由于腸道穩態下IL-22分泌減少后機體內存在其他代償機制。
綜上,本研究主要發現mTORC1功能缺失后導致ILC3分泌IL-22能力下降,鑒于IL-22在腸道抗微生物感染中的重要作用,提示mTORC1可能參與腸道穩態維持和抗感染的過程。后續我們將通過鼠類檸檬酸菌感染模型進一步驗證。另外,已有研究[25]發現mTOR依賴性的糖酵解等代謝程序調控了Th17細胞的增殖和效應功能。鑒于ILC3和Th17細胞之間的相似性,mTORC1對ILC3功能的調控是否也通過相關代謝通路仍需進一步探究。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中華詩詞(2022年6期)2022-12-31 06:41:24
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國科技論壇(2017年7期)2017-07-25 08:49:53
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
