組蛋白乙酰化酶對牙周膜干細胞內質網應激激活的調控研究*
2020-05-27 05:07:50賈婷婷張賢華
中華老年口腔醫學雜志 2020年2期
薛 芃 胡 楠 白 陽 蔡 川 賈婷婷 張賢華
慢性牙周炎發病機制和發展過程復雜。在慢性牙周炎長期炎癥微環境下,其內質網應激(endoplasmic reticulum stress,ERs)激活,未折疊蛋白反應(unfolded protein response,UPR)相關因子持續高表達,正常來源牙周膜干細胞(healthy periodontal ligament stem cells,H-PDLSCs)成骨分化能力持續降低。前期研究結果發現,短期炎癥微環境可導致ERs 和UPR 一過性升高[1,2]。因而猜想,長期和短期炎癥微環境對ERs 和UPR 的激活作用不同,可能是在長期炎癥微環境使細胞發生了轉錄水平或者轉錄后水平的調控。組蛋白乙酰化酶(histone acetyltransferases,HATs)可以在炎癥微環境影響下改變PDLSCs 轉錄后調控功能[3-5];HATs 還可影響ERs 表達[6-8]和干細胞分化功能[9,10]。因此,本研究通過篩選HATs 在炎癥微環境來源的PDLSCs(periodontitis periodontal ligament stem cells,P-PDLSCs)中的表達,繼而深入探究通過HATs 影響PDLSCs 中內質網應激持續性激活的可能作用機制。
1.材料和方法
1.1 主要儀器 YJ-875 型超凈工作臺(蘇州凈化設備廠);二氧化碳恒溫培養箱(Thermo,美國);6 孔板、平底96 孔板、底面積為25cm2培養瓶(Falcon,美國);體式顯微鏡、倒置相差顯微鏡及照相系統(Olympus,日本);透射電鏡(FEI,美國);CFX96 實時熒光定量PCR 儀(Bio-Rad,美國);酶聯免疫檢測儀(Thermo,美國);紫外線分光光度儀(Eppendorf,德國)。
1.2 主要實驗試劑 α-最低必需培養基(alphaminimum essential medium,α-MEM)、雙抗磷酸鹽緩沖液(phosphate buffered solution,PBS)、L-谷氨酰胺、青霉素及鏈霉素(Gibco,美國);胎牛血清(浙江天杭生物科技股份有限公司);胰蛋白酶(Amresco,美國);Ⅰ型膠原酶、維生素C、地塞米松、β-甘油磷酸鈉(Sigma,美國);TRIzol Reagent(Invitrogen,美國);實時定量PCR 試劑盒及cDNA 反轉錄試劑盒(Takara,日本)。
1.3 樣本收集與細胞分離培養 人正常來源牙周膜干細胞(H-PDLSCs):收集解放軍總醫院第一醫學中心口腔頜面外科門診30 至45 歲患者因正畸需要拔除、無齲的健康前磨牙或第三磨牙;人炎癥來源牙周膜干細胞(P-PDLSCs):收集解放軍總醫院第一醫學中心口腔頜面外科門診30 至45 歲患者因牙周炎需要拔除的牙齒。本項研究時間為2018年1 月至2018 年12 月。研究經解放軍總醫院倫理委員會批準,所有受試者均簽署了知情同意書。牙齒拔除后用含雙抗(青、鏈霉素)的PBS 液反復沖洗,用銳利刀片剝離牙根表面的牙周膜組織,剪成1mm3小塊接種于6 孔培養板內。每3 天換液1 次,待細胞從組織塊邊緣爬出并生長達80%匯合即原代牙周膜細胞;取原代牙周膜細胞,調整細胞密度至10~15 個/ml 接種于96 孔板(每孔0.1ml),37℃、CO2孵箱培養24h,標記單細胞孔后繼續培養;待形成克隆并達孔底面積1/3~1/2 時,胰酶消化,將多單克隆來源細胞懸液混合,體外擴增繼續培養。取第3 至5 代進行實驗。
1.4 實時定量PCR 檢測HATs 在P-PDLSCs中表達 經上述方法傳代的第4 代P-PDLSC 消化、離心收集后,將細胞密度調整至2×105個/ml,接種至6 孔板,24h 后終止培養;采用Trizol 試劑提取細胞總RNA,測定RNA 樣品濃度(A260/A280≥1.8);反轉錄合成cDNA,觀察HATs 相關基因的表達情況,PCR 引物序列見表1。本項研究所用引物序列由Primer Premier 5.0 設計,上海生工生物工程技術有限公司合成。取1.0μl 反轉錄合成的cDNA 在10μl 反應體系中進行實時PCR擴增。以甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH) 作為內參,通過2-ΔΔCt(ΔCt=Ct目的基因-Ctβ-GAPDH,ΔΔCt=ΔCt目的基因-ΔCt對照,Ct 為循環次數)法檢測各基因mRNA 相對表達量。每例樣品及陰性對照均設3 個平行復孔,取均值。
1.5 小干擾RNA(Small interfering RNA,siRNA)的轉染
1.5.1 轉染方法 5μl Lipofectamine 2000 溶于250μl 無血清無雙抗α-MEM 培養液制成A 液,室溫孵育5min;5μl siRNA 溶于250μl 無血清無雙抗α-MEM 培養液制成B 液;將A 液和B 液混合,室溫放置20min。吸取細胞培養液,用無血清無雙抗α-MEM 培養液清洗細胞一次,將1000μl 無血清無雙抗α-MEM 培養液加入A 液和B 液的混合液,混勻后加入細胞培養板;轉染6h 后,倒去含有轉染混合物的培養基,PBS 清洗,每孔加入細胞培養液2ml,繼續培養。
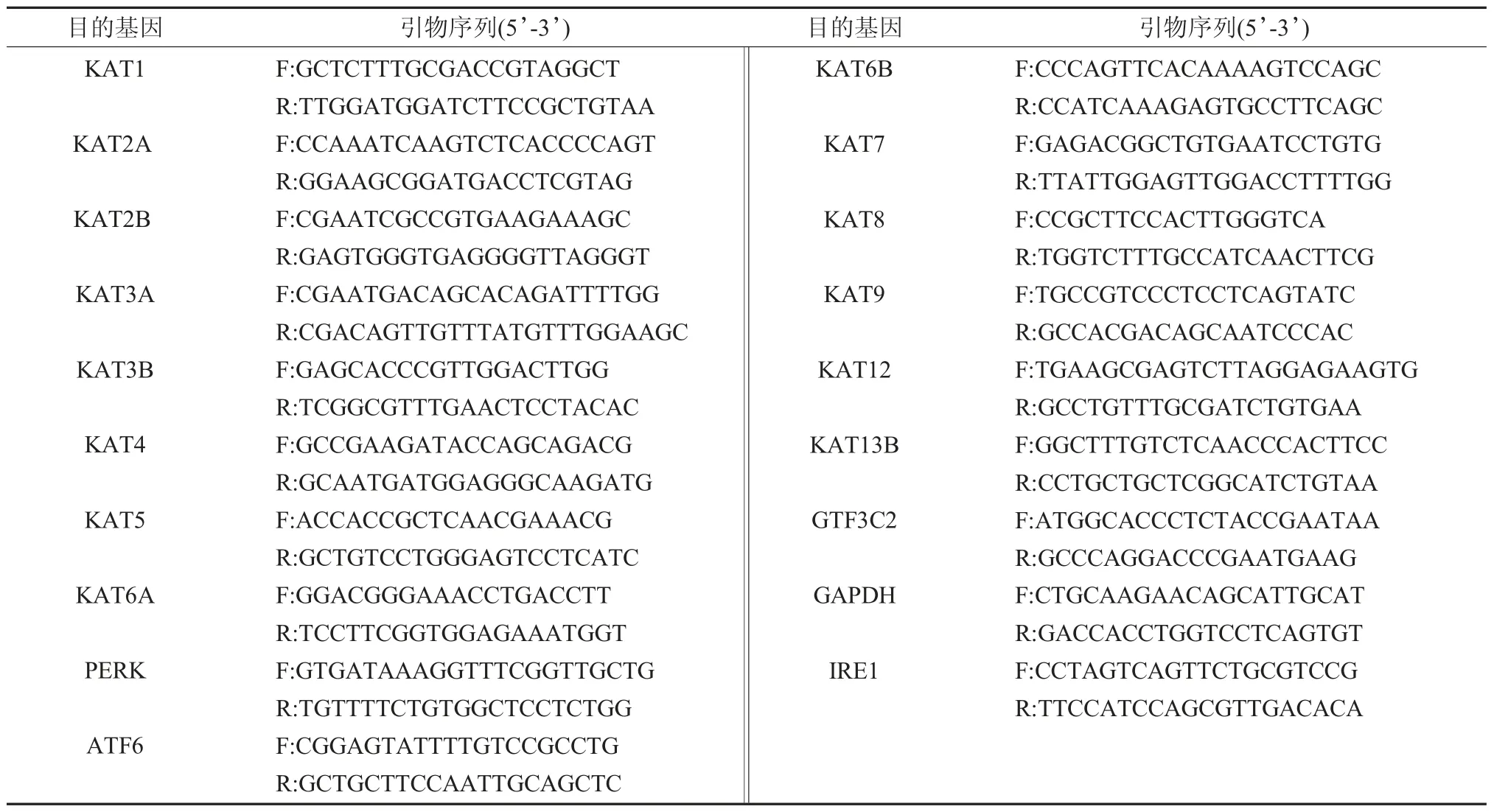
表1 本實驗所用實時定量PCR目的基因引物序列及長度
1.5.2 轉染效率檢測 轉染24h 后提取RNA,利用實時定量PCR 技術檢測siRNA 干擾效率在mRNA 水平的表達。每個轉染樣品設置三個復孔。
1.5.3 siKAT2A、siKAT3B、siKAT6A 和siKAT6B 后UPR 信號分子表達檢測 對H-PDLSCs進行siKAT2A、siKAT3B、siKAT6A 和siKAT6B轉染,以正常H-PDLSCs 為對照組。轉染24h 后提取RNA,利用實時定量PCR 技術檢測UPR 信號分子(PERK、IRE1、ATF6)mRNA 的相對表達量,PCR 引物序列見表1。
1.6 透射電鏡觀察siKAT6B 后H-PDLSCs中內質網形態變化H-PDLSCs 以2~3×107個/孔接種于六孔板24h 后,使用胰酶常規消化,800rpm 離心5min,棄上清,將PBS 沖洗后的樣品移入1.5 ml Ep 管,800rpm 離心5min,4℃固定12h,在室溫下進行脫水、滲透、聚合、切片,使用3%的醋酸鈾(30min)、枸櫞酸鉛(10min)雙染。透射電鏡觀察細胞內質網形態和大小,評估內質網形態的變化。
1.7 統計學方法 采用SPSS 17.0 統計軟件對結果數據行統計學分析,計量資料采用均數±標準差()表示,兩組間比較采用獨立樣本t 檢驗;以雙側P<0.05 為差異有統計學意義。
2.結果
2.1 篩選4 個HATs 在P-PDLSCs 中的表達變化 有限稀釋法克隆化培養PDLSCs 的鑒定見課題組之前已發表文章[1,11,12]。首先篩選了文獻報道與炎癥發生發展有密切關系,同時與內質網應激相關聯的HATs,見表2。結果顯示,與對照組比較,KAT2A、KAT3B、KAT6A 和KAT6B 4 個HATs在P-PDLSCs 中表達顯著降低,差異具有統計學意義(P<0.05)。
2.2 低表達KAT6B 可以激活UPR 中PERK通路 成功證明KAT2A siRNA、KAT3B siRNA和KAT6A siRNA、KAT6B siRNA 沉默有效,表3。同時,KAT6B siRNA 可以引起UPR 中PERK 信號分子mRNA 相對表達量的升高,而對其他UPR信號分子表達無明顯作用;而其他三個HATs siRNA 的使用并沒有使UPR 信號分子mRNA 的表達量升高,表4。
表2 HATs 在P-PDLSCs mRNA 相對表達量()
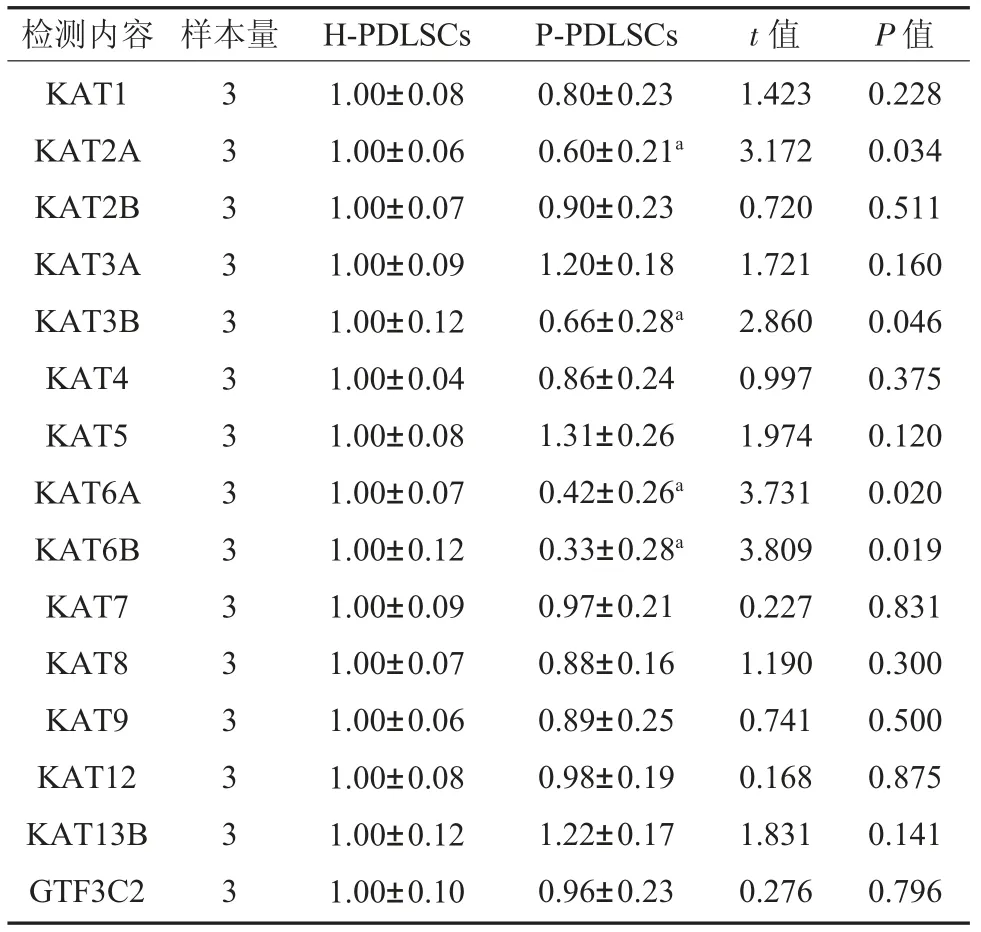
表2 HATs 在P-PDLSCs mRNA 相對表達量()
注:對照組:正常來源牙周膜干細胞;P-PDLSCs:炎癥來源牙周膜干細胞;KAT1、KAT2A、KAT2B、KAT9 為組蛋白乙酰轉移酶Gcn5/PCAF 家族成員;KAT5、KAT6A、KAT6B、KAT7、KAT8 為組蛋白乙酰轉移酶MYST 家族成員;KAT3A、KAT3B 為組蛋白乙酰轉移酶p300/CBP 家族成員;KAT4 為組蛋白乙酰轉移酶TAF 家族成員;KAT12、KAT13B、GTF3C2 為組蛋白乙酰轉移酶核受體共激活因子家族成員;a 表示與對照組相比P<0.05。
表3 4 個HATs siRNA mRNA 相對表達量()
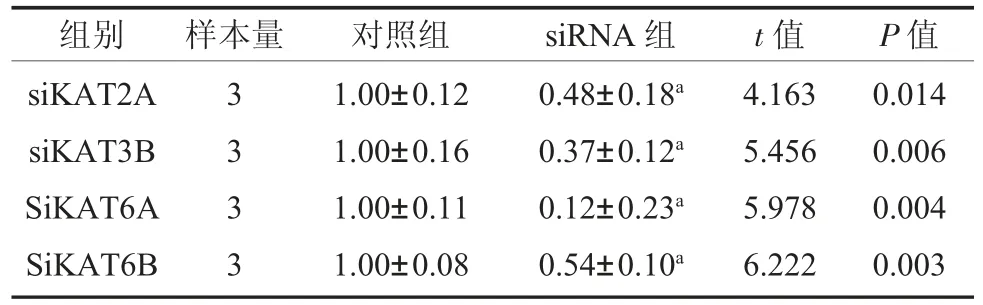
表3 4 個HATs siRNA mRNA 相對表達量()
注:對照組:正常來源牙周膜干細胞;KAT2A 為組蛋白乙酰轉移酶Gcn5/PCAF 家族成員;KAT3B 為組蛋白乙酰轉移酶p300/CBP家族成員;KAT6A、KAT6B 為組蛋白乙酰轉移酶MYST 家族成員;a 表示與對照組相比P<0.05。
2.3 低表達KAT6B 后H-PDLSCs 中內質網形態變化 我們利用透射電鏡技術觀察在H-PDLSCs中使用KAT6B siRNA 后內質網形態的變化。結果顯示,使用KAT6B siRNA 后,其內質網管腔呈現異常擴張和腫脹,且粗面內質網核糖體數量降低,圖1。
表4 4 個HATs siRNA 對UPR關鍵信號分子作用mRNA 相對表達量()
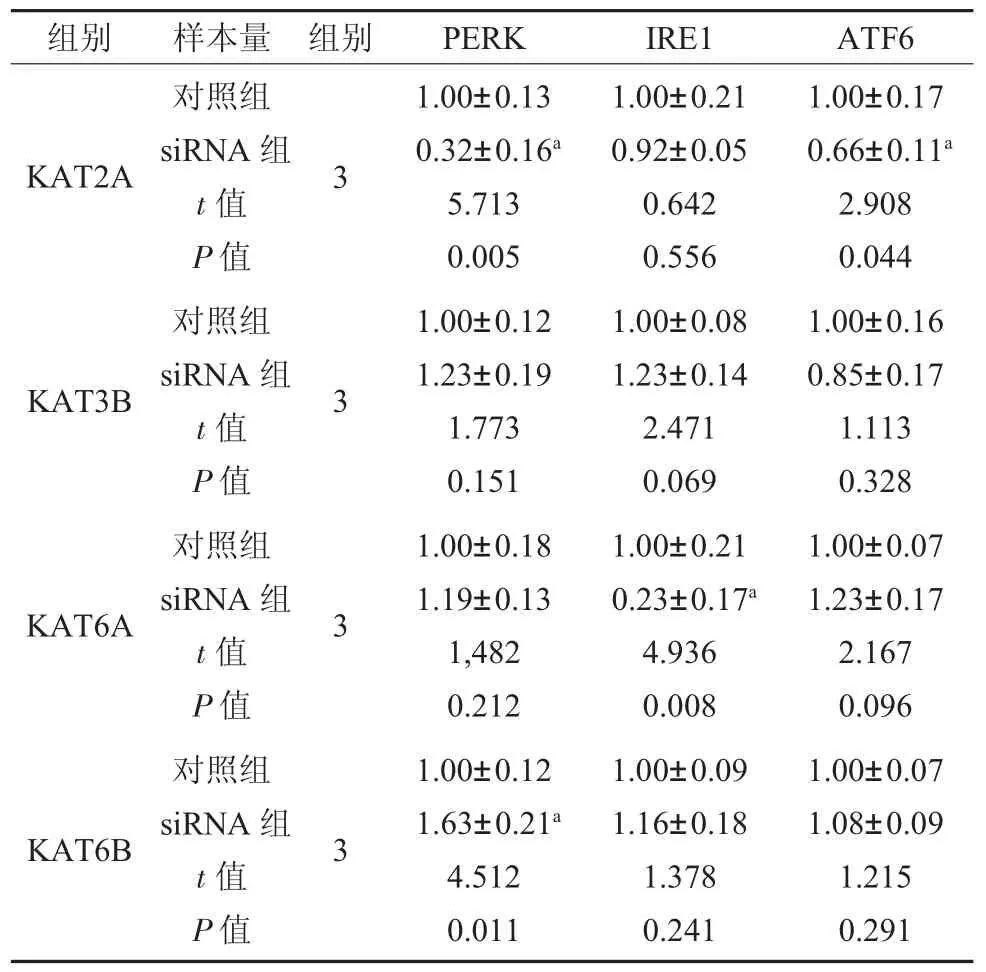
表4 4 個HATs siRNA 對UPR關鍵信號分子作用mRNA 相對表達量()
注:對照組:正常來源牙周膜干細胞;KAT2A 為組蛋白乙酰轉移酶Gcn5/PCAF 家族成員;KAT3B 為組蛋白乙酰轉移酶p300/CBP家族成員;KAT6A、KAT6B 為組蛋白乙酰轉移酶MYST 家族成員;PERK 為蛋白激酶受體樣內質網激酶;IRE1 為肌醇需要酶1;ATF6 為活化轉錄因子6;a 表示與對照組相比P<0.05。
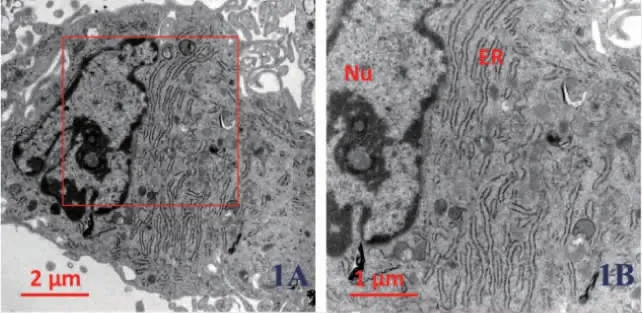
圖1 正常牙周膜干細胞(H-PDLSCs)內質網形態的透射電鏡觀察
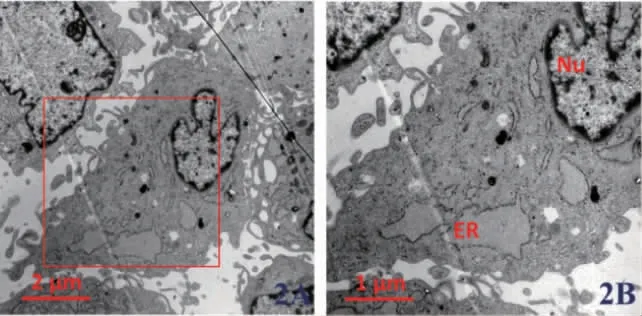
圖2 低表達KAT6B 后正常牙周膜干細胞(H-PDLSCs)內質網形態的透射電鏡觀察
3.討論
慢性牙周炎是以細菌感染為主的多因素進展性疾病,在其臨床表現中,以牙槽骨喪失最為嚴重[13]。近年來越來越多的研究表明,細胞在受到缺血、低糖、低氧和感染等外界刺激時,其蛋白折疊和降解等程序將受到干擾,產生ERs,為了適應這種應激反應,細胞通過UPR 對細胞進行保護和調節。UPR是一個復雜和協調并存的細胞應答反應,它通過內質網的三種跨膜受體完成調控:蛋白激酶樣內質網激酶(Protein kinase receptor like ER kinase,PERK)、轉錄激活因子6(Activating transcription factor 6,ATF6)和肌醇需要酶1(Inositol requiring enzyme1,IRE1)、也稱作ERN1。細胞處于靜息狀態下,三種內質網跨膜蛋白與GRP78 穩定結合,保持非活性狀態;細胞處于應激狀態下,非折疊蛋白積聚,三種跨膜蛋白與GRP78 的解離,產生UPR。因此,UPR 的目的是減少非折疊蛋白的積壓以及恢復內質網正常功能,使細胞正常存活[14]。如果應激不能有效緩解,這個保護性的信號通路就會轉向促凋亡反應。本課題組首次發現ERs 在炎癥來源的PDLSCs 中表達激活,且可通過UPR 的PERK 通路導致PDLSCs 成骨分化下降,證明炎癥微環境下ERs 的產生是調控干細胞成骨分化的重要原因[1,2]。但炎癥微環境下ERs 激活后如何持續影響炎癥的進程尚不清楚。
近年來越來越多研究表明,慢性牙周炎與表觀遺傳修飾有關。表觀遺傳主要包括DNA 甲基化、微小RNA(micro RNA,mi RNA)修飾、組蛋白乙酰化和長鏈非編碼RNA(long noncoding RNA,lnc RNA)修飾等[15]。組蛋白乙酰化屬于表觀遺傳修飾其中一種,當組蛋白結構發生改變,影響基因轉錄。目前表觀遺傳與牙周炎的發生在組蛋白乙酰化領域的研究較為表淺,因此,本實驗首先通過對比正常及牙周炎癥來源的PDLSCs 中HATs 的表達水平,發現與H-PDLSCs 相比,P-PDLSCs 中KAT2A、KAT3B、KAT6A 和KAT6B 四個基因表達出現顯著下降。此研究表明,HATs 與牙周炎發生發展具有明顯相關,炎癥微環境可以引起4 個HATs 的低表達。
為進一步明確篩選出的4 個低表達的HATs和內質網應激的激活的關系,我們利用siRNA 分別對4 個HATs 進行抑制,同時檢測UPR 信號分子(PERK、IRE1、ATF6)mRNA 相對表達量。我們發現,只有低表達KAT6B 后可以導致PDLSCs中UPR 信號分子激活,而其他3 個HATs 低表達后,不能引起UPR 激活。同時,我們使用透射電鏡直觀觀察到siKAT6B 后,內質網腔擴張,作為ERs 和UPR 被激活的標志[16]。這些實驗結果顯示,KAT6B 在對內質網應激和UPR 的激活中發揮了重要作用。
KAT6B 屬于組蛋白乙酰轉移家族中MYST 家族的成員,它結構包括NEMM(N 端Enok)結構域,1 段催化MYST 模塊,2 段串聯的鋅指結構(Plant homeodomain,PHD)和1 段富含谷氨酸鹽/天冬氨酸鹽(Glutamate/aspartate-rich,ED)結構域和富含絲/蛋氨酸(Serine/methionine,SM)結構域[17-19]。KAT6B 的MYST 結構域具有乙酰化活性,可以乙酰化組蛋白H3 的賴氨酸9 和14;KAT6B的C 端結構域是富含SM 的酸性區域,它們存在潛在的轉錄激活活性,但對于這一區域的功能相應的報道較少。比如,KAT6B 與DNA 不直接結合,它們作為DNA 結合的轉錄激活因子的共調解因子存在[17];還有研究表明,KAT6B 可以介導RUNX轉錄因子的激活[17,20,21]。
綜上所述,本項研究結果顯示,牙周炎癥微環境可以導致組蛋白乙酰化酶KAT6B 降低,同時導致ERs 和UPR 激活,但KAT6B 對UPR 持續激活的具體機制尚待深入研究。
