不同生長調節劑對‘新球蜜荔’體胚發生的影響及其組織學觀察①
2020-05-22 12:48:32李煥苓王樹軍孫進華王家保
熱帶農業科學 2020年2期
王 果 李煥苓 王樹軍 孫進華 李 芳 王家保
(中國熱帶農業科學院環境與植物保護研究所/農業農村部儋州農業環境科學觀測試驗站 海南海口571737)
荔枝(Litchi chinensisSonn.)屬無患子科荔枝屬常綠喬木,是我國南方重要的經濟果樹之一[1]。產業中的病危害嚴重、采后損失較大、成花不穩等問題迫切需要優良品種。傳統的育種效率已經難以滿足目前市場的需求,生物技術育種時效高成為當前植物品種改良的主要手段,已經成功應用于香蕉[2]、番木瓜[3]、番茄[4]、苜蓿[5]、水稻[6]、蘋果[7]等植物品種改良中。建立高效穩定離體再生技術體系是植物生物技術育種的基石,體胚發生是建立木本植物離體再生的主要途徑。體胚具有兩極性結構,可以直接形成再生植株,在無性繁殖上具有較大優越性[8]。開展荔枝體胚發生研究,對于植物基因工程、細胞工程、雜交育種、良種快繁、轉基因受體和突變體篩選等具有重大價值,同時有助于研究細胞離體培養中形態發生、細胞分化與胚胎發生等重大理論問題。
荔枝品種不同,其離體再生體系建立難易不同,體胚高頻發生是建立荔枝離體再生技術體系的基礎。目前僅有15 個荔枝品種獲得再生苗[9],其離體再生體系不穩定,再生率較低,最基本的原因在于未能建立高頻穩定的體胚發生體系從而獲得大量體胚。‘新球蜜荔’是海南原產的優異荔枝資源品種,果實外觀好,風味佳,產量好;生長量小,栽培管理不復雜,建立‘新球蜜荔’離體再生技術體系,可為加快其在育種上的改良及生產推廣應用提供理論基礎。本研究擬以‘新球蜜荔’花藥為外植體獲得的胚性愈傷組織為材料,研究不同生長調節劑對‘新球蜜荔’體胚誘導的影響,建立體胚成熟及萌發體系,同時對體胚發生過程中的組織形態學觀察,揭示荔枝體胚發生和發育過程,為果樹體胚發生研究提供基礎。
1 材料與方法
1.1 材料
‘新球蜜荔’花藥誘導保持所獲胚性愈傷組織為材料,比較不同生長調節劑對‘新球蜜荔’體胚高頻發生的影響。
1.2 方法
1.2.1 體胚發生
將繼代保持18 d 旺盛生長的‘新球蜜荔’胚性愈傷組織接種到添加不同生長調節劑的MS 培養基上(表1)誘導體胚發生。每瓶接種約0.1 g,每個處理各接種10 瓶;接種后置于(25±2)℃環境中黑暗培養;以10瓶為一重復,實驗獨立進行3次;約7 周時統計體胚個數,以直徑≥0.3 cm 的體胚為準。
1.2.2 體胚成熟
參考王果[20],體胚形態多樣,有雙子葉胚、團狀胚、喇叭胚等9種形態,依據其顏色可分為乳白胚和透明胚,乳白胚即體胚乳白色,經過成熟后有進一步萌發的潛力。透明胚為淡黃色透明胚,在成熟過程中逐漸褐變死亡,不能萌發。
將誘導53 d、直徑≥0.3 cm 的‘新球蜜荔’乳白胚與透明胚分別轉接到不同濃度ABA 與PEG-600組合的成熟培養基上,每處理約接種30 個,實驗獨立進行2 次;約65 d 時觀察并記錄體胚成熟情況。
1.2.3 體胚萌發
將‘新球蜜荔’成熟胚轉接到萌發培養基上,對比不同成熟培養基上成熟體胚的萌發效果。每瓶接種約4個體胚,每個處理各接種20瓶;約30 d統計體胚萌發量,觀察并記錄體胚萌發過程。
1.2.4 愈傷組織形態學觀察
取‘新球蜜荔’T3、T7、T11 體胚培養基上培養3、6、9、12、15、18、21、24、27 d 培養物進行組織形態學觀察。
2 結果與分析
2.1 NAA對體胚發生的影響
NAA 0.1 mg/L與KT 5 mg/L組合中,‘新球蜜荔’體胚發生率、發生數量、乳白胚比例都較未添加NAA 培養基上的明顯降低。分別添加NAA 0.1 mg/L與ZT 5 mg/L、TDZ 0.5 mg/L 的兩個組合中,‘新球蜜荔’體胚發生率、發生數量、乳白胚比例都比未添加NAA的高。
2.2 不同生長調節劑組合對3 個荔枝品種體胚發生的影響
從圖1 與表1 可以看出,不同種類、不同濃度的生長調節劑(ZT、KT、TDZ)對‘新球蜜荔’體胚發生影響不同。影響較大的是TDZ,其次依次是ZT 與KT;其中添加TDZ 的培養基體胚發生率達100%,體胚發生數量最多,為6 010 個/g FW;添加ZT 的培養基體胚發生率均為100%,體胚數量最多達5 540 個/g FW;添加KT 的培養基體胚發生率最高只有90%,體胚數量為4 110 個/g FW。添加一定濃度ZT、KT 組合的培養基均發生體胚,數量最高達5 690個/g FW。
不同種類、不同濃度生長調節劑對‘新球蜜荔’體胚發生數量質量影響都不同。添加TDZ 0.5 mg/L 培養基上體胚發生數量最多,達6 010 個/g FW,乳白色體胚占93.29%,多為球形胚與細長子葉胚,直徑多在0.3~0.7 cm,有部分乳管胚直徑達1.7 cm;TDZ 濃度升高或降低,其體胚數量都急劇減少,且透明胚比例升高,在TDZ 0.1 mg/L時全部是透明胚。在添加不同濃度ZT 的培養基上,以添加5 mg/L ZT 培養基上的白色體胚發生率最高,乳白色體胚多為球形,不黏連,一碰即落,不似TDZ 中的球形胚成塊;體胚整體在0.3~0.6 cm,體積較TDZ、KT 培養基上的大小均一,無明顯差距;體胚數量較TDZ 0.5 mg/L培養基上的體胚數量少,隨著TDZ 濃度升高或降低,其體胚數量都減少,分化大量透明胚,透明胚體積相對較大。在添加不同濃度KT的培養基上,體胚發生數量較少,最高僅4 110個/g FW;乳白色體胚比例也較低,為1.95%;隨著KT濃度升高,其體胚發生率與白色體胚比例也增大,添加7 mg/L KT,體胚發生數量達3 810 個/g FW,沒有僅添加5 mg/LKT 培養基上的體胚數量多,但是乳白色體胚比例較其微高;添加KT 培養基上的胚性愈傷組織全部褐變。添加ZT 3 mg/L 與KT 2 mg/L 組合的培養基上體胚發生數量達5 690 個/g FW,乳白色體胚占89.7%;該培養基上體胚全分化,透明胚相對大,0.4 cm以上;且還分化較多0.1 cm 球形、小乳白胚;子葉胚彎曲,0.3 cm 且漸成熟轉綠。但同時添加ZT2 mg/L、KT 3 mg/L 培養基上的體胚則較多分化透明胚,體積形態不規則,可分化至子葉胚,球形胚較少。
由此可見,過高或過低生長調節劑對荔枝體胚發生都是不利的。確定‘新球蜜荔’體胚高頻發生的適宜培養基為添加TDZ 0.5 mg/L、NAA 0.1 mg/L的MS培養基。
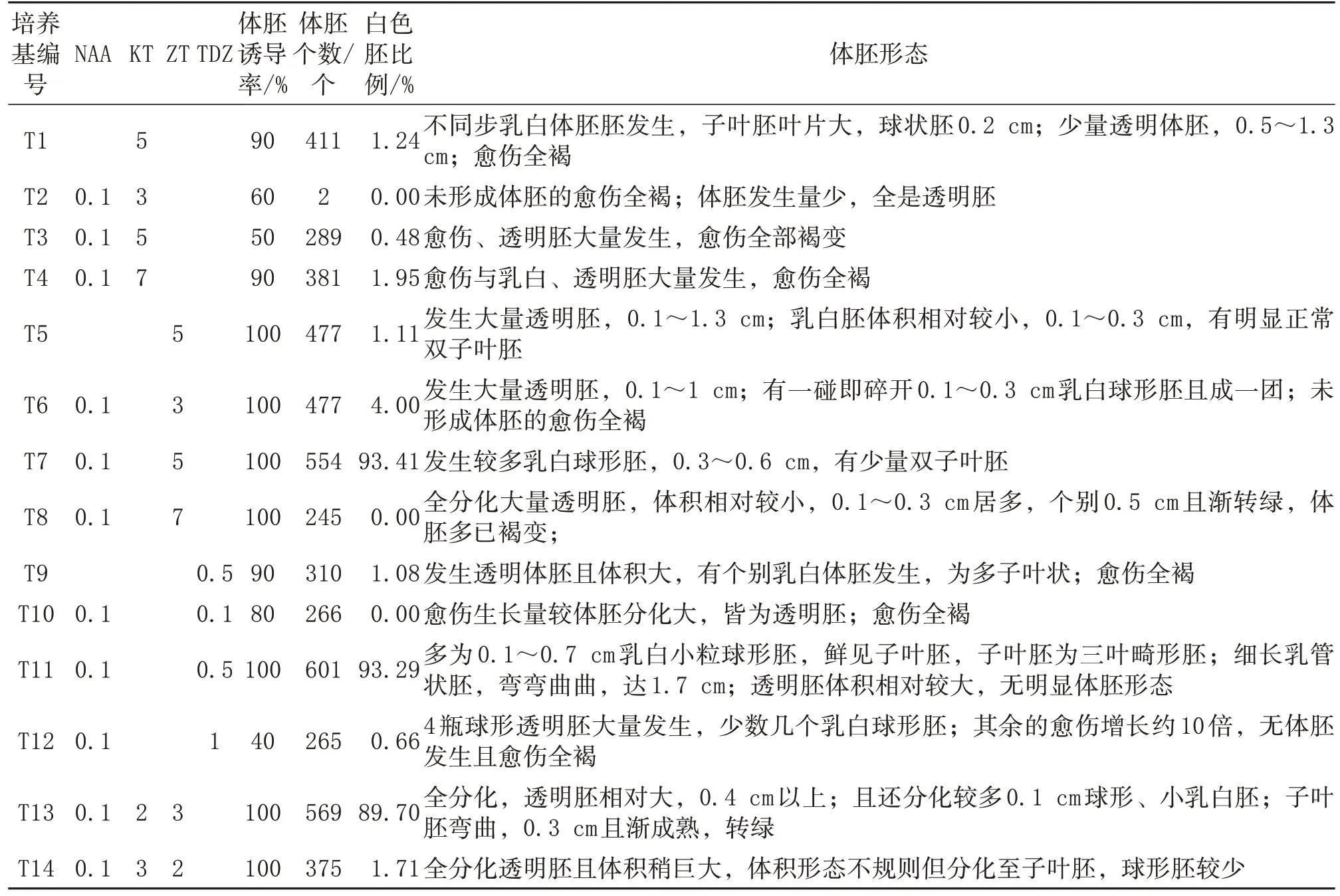
表1 不同生長調節劑對‘新球蜜荔’體胚發生的影響
2.3 ABA 及PEG 不同濃度組合對體胚成熟及萌發的影響
2.3.1 對體胚成熟的影響
無論是否添加ABA,體胚褐死率都隨著PEG-600濃度升高出現先降低后升高;而當無添加PEG-600 時,褐死率隨著ABA 濃度升高出現先升后降,最低達42.22%;當ABA 濃度為1 mg/L 時,褐死率明顯上升;當ABA濃度為2 mg/L 時,褐死率微降后升。因此,僅添加PEG-600 或1 mg/L ABA 都能明顯降低體胚褐死率,但同時添加兩者其褐死率反而較單獨添加的高;不過,高濃度的ABA(2 mg/L)與高濃度PEG-600(50 g/L)組合其褐死率反而降低,接近單獨添加PEG-600的褐死率。
PEG-600 通過提高培養基滲透勢對體胚含水量起作用,而ABA 能改善體胚質量,添加了ABA 的培養基處理乳白色體胚都較鮮白,飽滿。因此選擇PEG-600 25 g/L 促進體胚成熟,在體胚的特定或某一階段選擇1 mg/L ABA來調控體胚狀態。在上述不同體胚成熟培養基上,透明胚逐漸褐變死亡,個別乳白胚褐變(表2)。

表2 不同濃度ABA及PEG組合對‘新球蜜荔’體胚成熟的影響
2.3.2 對體胚萌發的影響
不同來源成熟體胚在萌發培養基上表現不同。來源于有添加PEG-600的成熟體胚褐死較多。當無PEG-600存在時,添加1 g/L ABA培養基上的體胚褐死最少;添加不同濃度PEG-600 后,ABA 對體胚褐死影響不是很大;而同時添加高濃度PEG-600 及ABA 后,體胚大多褐變;因此體胚成熟時單獨添加ABA對體胚萌發有促進作用。
無添加ABA 時,低濃度PEG-600(25 g/L)對體胚萌發有抑制作用,隨著濃度升高,體胚較少褐變;而當添加ABA 后,隨著PEG-600 濃度升高,其褐變程度加大。因此體胚成熟時單獨添加PEG-600對體胚萌發有促進作用。
體胚萌發過程中全部變綠,僅有一個體胚抽出莖段,但無真葉展開。個別乳白胚再生出鮮乳白色次生胚,體積較小,約0.5~0.8 cm,該種體胚在光下不能變綠;透明胚在萌發過程中逐漸褐死。在萌發培養基上有少數類似于莖、芽,或彎曲或泡狀化的畸形結構,有1 個體胚有不定根生成。
2.4 ‘新球蜜荔’體胚發生過程中的組織形態學變化
‘新球蜜荔’胚性愈傷組織細胞有橢圓形與圓形,細胞緊湊,內含物豐富,可見細胞質、細胞核及淀粉粒(圖1 b 紅色箭頭所指)。胚性愈傷組織中夾雜非胚性細胞,非胚性細胞體積較大,細胞核質比低,在體胚分化過程中逐漸裂解凋亡。胚性愈傷組織內部或者表面可見大小不一的單個或兩個以上相對孤立的胚性細胞或胚性細胞團,由此可推測胚性細胞可能起源于單個細胞,但不排除多個細胞的體胚發生方式;體胚發生的起源方式則同時有外起源(圖1 b 黃色箭頭所指)和內起源(圖1b 紅色、紫色箭頭所指)兩種。
胚性細胞體積小,核大,細胞質稠密,染色深,經過對稱分裂形成胚性細胞團。隨著培養時間延長,胚性細胞團形成明顯凸起或長條形原胚(圖1b 黃色箭頭所指),經過不對稱分裂,原胚表現出極強烈的極性,胚頂端下凹兩邊凸起形成子葉原基,分化為心形胚;子葉原基接著分裂分化,子葉伸長,發育成為魚雷形胚。魚雷形胚子葉繼續伸長,根冠細胞裂解(圖2 KT-18 d),形成具有明顯頂端分生組織、原形成層帶、發育良好的子葉和根的子葉胚。體胚在形成過程中或與愈傷組織緊密相連,或在不同發育階段時從愈傷組織中游離開來,有的體胚則互相黏連,而形成連體胚(圖2TDZ-12 d)。
在不同體胚發生培養基上,體胚發生時間不一致。ZT、TDZ 培養基上的愈傷組織在初期(ZT-3 d,TDZ-3 d)已經有原胚發生,ZT 培養基上,至9 d 時有多個原胚,12 d 時有心形胚及子葉胚出現,且12~15 d間出現大量心形胚及子葉胚;21~24 d主要是球形胚與魚雷形胚,至24 d 時多子葉胚居多。TDZ 體胚發生培養基上,12 d 時發現心形胚,15 d 時有大量心形胚出現,21 與24 d 出現大量的魚雷形胚及子葉胚,此時間是體胚開始大量形成子葉胚的階段。
KT 體胚發生培養基上,9 d 天時才隱約可見球形胚(圖2 KT-9 d),12 d 時球形胚大量發生,15~18 d時主要是球形胚及心形胚發生,21 d有子葉 胚出現,24 d子葉胚增多。
3 討論
3.1 不同生長調節劑種類及水平對體胚誘導的影響
除基因型外,激素種類及水平是影響植物離體再生的主要因素。KT、ZT 是果樹體胚發生最常用的植物生長調節劑,使用濃度范圍較大,在0.25~5 mg/L。橡膠[21]、荔枝[22]及芒果[23]等果樹上使用的KT 濃度較高,分別為1、3、5 mg/L,從而獲得大量體胚。雜交鵝掌楸[24]采用富含0.5 mg/L KT 或ZT 4 mg/L 獲得大量原胚。TDZ 常用于頑拗性植物體胚發生,其作用較強,使用濃度范圍較低。紅姜花[25]采用0.15 mg/L TDZ 誘導體胚,體胚誘導率高達4500 個/mL;當TDZ 為0.003 mg/L時,杉木[26]體胚誘導率最高;尾巨桉[27]胚性愈傷組織在附加NAA 0.1 mg/L 與TDZ 0.1 mg/L 的MS 培養基上得到體胚。本試驗通過添加NAA與不同濃度的細胞分裂素(KT、ZT、TDZ)對‘新球蜜荔’胚性愈傷組織體胚發生影響的比較發現,NAA、KT、ZT、TDZ對體胚發生均有作用,在添加TDZ 0.5 mg/L、NAA 0.1 mg/L MS 培養基上,體胚發生數量達6 010個/g FW。隨著激素濃度降低或升高,體胚發生數量都降低。適宜濃度KT、ZT組合對‘新球蜜荔’體胚發生相對也較好,體胚發生數量達5 690 個/g FW。
3.2 體胚成熟及萌發
ABA 是促進體胚成熟最有效的植物生長調節劑,常與滲透劑PEG用于促進體胚成熟。幾乎所有針葉樹都采用ABA 及PEG 組合來促進體胚成熟[28],其組合ABA常用濃度為10~50 mg/L,PEG常用PEG-8000,使用濃度范圍較大,從20~200 g/L。在培養基中添加0.5 g/L ABA 及PEG 可顯著提高栓皮櫟體胚成熟[32]。香榧體胚成熟處理中PEG-6000 40~80 g/L、ABA10 mg/L 比 較 適 合 體 胚 成 熟[33]。在‘新球蜜荔’體胚成熟過程中,25 g/LPEG-600有一定促進作用,ABA 1 mg/L 能改善體胚質量。體胚萌發時,僅有一個體胚抽出莖段。因對成熟體胚形態并無統一標準,均以成熟體胚具有較高萌發率作為判斷體胚是否發育成熟的依據,很多研報道提出了可以促進體胚成熟發育的因素但缺乏對其促進機理的研究,因此探明體胚發生機理才能突破體胚成熟發育不完全,萌發低的瓶頸。
3.3 體胚發生過程中的組織形態學變化
植物體胚發生重演合子胚發生途徑,但體胚發生部位及發生方式隨植物種類及生長調節劑作用而異,大部分植物體胚發生都是單細胞起源。王天地[34]研究發現,橡膠體胚發生部位同時存在于愈傷組織表層及愈傷組織內部的單個胚性細胞;Shirin[35]研究發現,疏花火燒蘭早期體胚起源于愈傷組織表層多個細胞團結構;Shen[36]發現,萬年青體胚來源于愈傷組織表層的單個胚性細胞。荔枝體胚發生過程中的組織形態學變化已有研究報道。鄧朝軍[14]指出,三月紅荔枝體胚發生與合子胚發生途徑一致;吉訓志[37]對比了2 個荔枝品種不同荔枝品種體胚發生初期愈傷組織的形態變化,指出愈傷組織形態不同,其體胚發生能力不同。王果[20]指出,‘妃子笑’荔枝愈傷組織形態不一,其體胚發生起源于愈傷組織內部單個胚性細胞,重演合子胚發生過程。本研究發現,‘新球蜜荔’體胚發生部位既有內部又有外部兩個部位,都是單細胞起源。通過調整胚性愈傷組織狀態來改善‘新球蜜荔’體胚形態多樣且高度不同步化,體胚與愈傷組織黏連或其游離時間早晚與體胚成團是否存在關系等有待于進一步研究。
