系統性紅斑狼瘡致病基因研究進展*
2020-05-18 10:51:14翟建昭孟妍明綜述李益洲武永康審校
國際檢驗醫學雜志 2020年9期
關鍵詞:信號
朱 盈,翟建昭,羅 娟,孟妍明 綜述,李益洲,武永康& 審校
(1.四川大學華西醫院實驗醫學科,四川成都 610041;2.四川大學化學學院,四川成都 610061;3.四川大學網絡空間安全學院,四川成都 610061)
系統性紅斑狼瘡(SLE)是一種多基因相關的自身免疫性疾病,表現出明顯的家族聚集性,且同卵雙生子的發病率一致性(24%)高于異卵雙生子(2%),基因研究對于SLE致病機制的研究十分重要[1]。SLE作為一種自身免疫性疾病,發病過程涉及固有免疫和適應性免疫,根據免疫反應過程可分為抗原識別、淋巴細胞間信號傳遞和免疫過程調節等(圖1),任何過程的錯誤或紊亂都可能增加SLE易感性。本文對SLE各免疫反應階段的主要致病基因進行綜述,以期為SLE遺傳與免疫信號通路的全面認識提供依據。
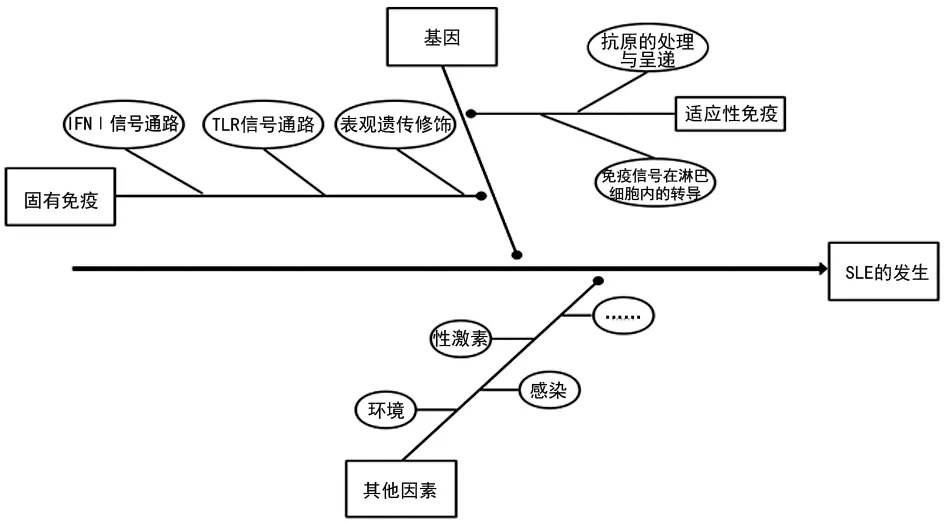
注:IFNⅠ表示Ⅰ型干擾素;TLR表示Toll樣受體。
圖1 SLE致病因素及發生過程
1 固有免疫應答
1.1IFNⅠ信號轉導途徑 IFNⅠ信號轉導途徑在抗腫瘤、抗病毒和免疫調節中都起著重要作用,是固有免疫應答的主要組成部分,在機體控制并清除病原體的過程中扮演著重要角色。免疫細胞通過模式識別受體接受抗原刺激后釋放IFNⅠ,IFNⅠ通路過度活化是SLE發病的一大原因。
1.1.1Janus激酶-信號轉導及轉錄激活因子(JAK-STAT)通路 STAT是干擾素依賴性基因表達的主要成分,JAK-STAT通路過程涉及多種細胞因子、激素和生長因子等的信號轉導,介導細胞正常增殖、分化、活化、成熟等,而異常的信號轉導則會導致SLE等多種自身免疫性疾病。STAT4作為STAT家族的重要分子,主要表達于淋巴細胞、巨噬細胞和樹突狀細胞等抗原呈遞細胞(APC)的細胞質,能在膜結合受體的刺激下活化進入細胞核,調節相關基因的轉錄和表達并且傳輸信號,該基因的多態性與SLE的發病呈強關聯,如rs7574865 T>G的突變可能與SLE有關[2]。
1.1.2TLR信號通路 TLR是一種跨膜蛋白,是固有免疫的重要組成部分,也是連接固有免疫與適應性免疫的橋梁。自身抗原與TLR結合后,可激活IRAK等基因,導致核因子κB(NF-κB)、干擾素調節因子(IRF)5和IRF7的活化,最終使細胞因子增加和自身抗體產生。
1.1.2.1白細胞介素(IL)-1相關受體激酶1(IRAK1) IRAK1是一種有效的NF-κB激活劑,位于X染色體上,與SLE存在強烈的相關性,可能是導致女性患病率較高的原因之一[3]。TLR與自身抗原結合后,將信號傳遞給下游的IRAK1。SLE患者CD4+T細胞內IRAK1轉錄水平顯著增加,并且與疾病活動度呈正相關[4]。使用IRAK1抑制劑可以抑制Th17細胞的分化,有望成為新的治療方案。除了轉錄水平增加以外,IRAK1(rs3027898)的多態性也會增加SLE的易感性。此外,IRAK1在激活NF-κB的過程中,受到非編碼微小RNA146a(miRNA146a)的靶向調節,因此,miRNA146a(rs2910164)的多態性也會影響SLE的發病[5]。
1.1.2.2干擾素調節因子家族(IRFs) IRFs是轉錄因子的一種,參與病毒相關的干擾素活化、細胞分化凋亡及免疫系統活性的調節。IRF作為IFN通路重要調控因子,在自身免疫反應發生的過程中起重要作用。研究證實IRF5基因rs729302 A等位基因與SLE的發生密切相關[6-7]。另外發現IRF是免疫應答的重要分子,如IRF5與細胞因子的產生有關,并且能夠誘導IFNⅠ的生成;IRF7可以影響IFNⅠ信號通路,與SLE發病密切相關[8]。
1.1.3表觀遺傳修飾
1.1.3.1miRNA miRNA,即小的非編碼RNA,通過抑制mRNA的轉錄和翻譯對靶基因進行表觀修飾,這類分子對SLE發病起著重要的作用。各類miRNA作用復雜,如miRNA-146a和miRNA-155能干擾TLR-7和TLR-9下游的細胞內信號傳導途徑,并影響漿細胞樣樹突狀細胞中IFNⅠ的生成。在T細胞和B細胞中,miRNA-126、miRNA-21、miRNA-146a、miRNA-155和miRNA-1246等可能通過表觀遺傳修飾影響基因表達[9]。有研究表明SLE患者體內miRNA-146低表達,導致其靶基因TRAF6和IRAK-1表達上升,引起IFN通路激活,刺激SLE患者疾病發展[10]。
1.1.3.2甲基化修飾 DNA低甲基化在SLE的發病過程中起著非常重要的作用,DNA的低甲基化會導致一些自身免疫疾病相關基因過度表達,如CD11b、CD40L等[11],從而導致自身免疫性疾病的發展。DNA甲基化與淋巴細胞的發育有關,SLE患者的幼稚T細胞呈現整體低甲基化[12],此外還會影響IL-2、IL-4和IFN-γ等細胞因子的表達,實際上多種表觀遺傳修飾往往是同時發生、共同作用的。
1.2NF-κB信號活化途徑 NF-κB通路是體內多種信號通路的交叉點,多種受體的激活都可以導致其NF-κB的活化,如TLR、B細胞受體(BCR)等。NF-κB信號通路在體內起重要的信號轉導作用,其調節異常會引起包括SLE在內的自身免疫性疾病,在這條通路涉及的基因中,有許多與SLE的發生有關。
1.2.1腫瘤壞死因子誘導蛋白3(TNFAIP3) TNFAIP3可被腫瘤壞死因子(TNF)誘導表達,在體內抑制NF-κB通路的活化及TNF介導的細胞凋亡。已有研究證實TNFAIP3 rs2230926多態性與SLE之間的關聯,在中國人群中也已經報道過,該位點的改變使對NF-κB的抑制減弱,從而增加SLE的易感性[13]。
1.2.2TNFAIP3相互作用蛋白1(TNIP1) TNIP1編碼的產物與TNFAIP3類似,二者協同抑制NF-κB信號傳導途徑,當兩者上調表達時,NF-κB失調,極易導致SLE及狼瘡性腎炎等自身免疫性疾病的發生,該基因rs10036748位點也是一個被多次證實的SLE強關聯位點[14],其多態性也會使得抑制作用減弱。
1.2.3整合素亞基α(ITGAM) ITGAM基因編碼CD11b,與單核-巨噬細胞系統相關,也涉及粒細胞的黏附趨化作用和攝取補體包被的顆粒和病原體的作用,可以調節固有免疫過程和包括NF-κB信號通路在內的一些信號傳導途徑。ITGAM的遺傳變異與SLE、狼瘡性腎炎和其他SLE相關并發癥的易感性密切相關[15]。目前較常見的與SLE相關的由ITGAM基因編碼的CD11b突變單核苷酸多態性(SNP)有rs11574637和rs1143679,其多態性均會增加SLE的易感性[16-17]。
2 適應性免疫應答
2.1自身抗原處理與提呈
2.1.1主要組織相容性復合體(MHC)Ⅱ類基因 MHC是最早被報道的與SLE易感性相關的基因,人類MHC基因編碼產物通常被稱為人白細胞抗原(HLA),其中MHCⅡ類分子可以識別外源性抗原,主要在抗原呈遞細胞表面表達,在免疫應答的始動階段將經過處理的抗原片段遞呈給CD4+T細胞。HLA是目前發現的與自身免疫性疾病遺傳性有最強關聯的基因。已有多種研究表明,屬于MHCⅡ類的HLA-DR2和HLA-DR3的單倍型與SLE易感性相關,是SLE的強烈風險因素[18-19]。HLA-DQA2的SNP(rs2301271)與SLE有顯著相關性,并且在rs2187668處觀察到抗dsDNA和HLA-DR3之間的強關聯,進一步支持HLA區域在SLE致病因素中的重要性[20]。
2.1.2共刺激分子對
2.1.2.1OX-40L與OX40 腫瘤壞死因子超家族成員4(TNFSF4)基因又稱為OX-40L,主要在活化的抗原呈遞細胞和血管內皮細胞上表達,其受體TNFRSF4(也稱為OX40)主要表達在CD4+T細胞表面,已有大樣本研究顯示,TNFSF4的多個位點多態性均與SLE的發病相關,在中國內地漢族人群證實rs1234315多態性與SLE的發病強相關[21]。
2.1.2.2分化簇(CD)80與CD28 CD80分子表達在抗原呈遞細胞表面,可以與T細胞表面的CD28分子結合,活化膜受體從而刺激T細胞增殖分化,在SLE患者體內CD80+細胞顯著增加。編碼CD80的 rs2222631和rs6804441的基因多態性與SLE顯著相關,且彼此獨立[22]。此外,在一項SLE小鼠模型實驗中,阻斷CD80與CD28之間的共刺激作用可以有效阻止狼瘡性腎炎的發生[23]。
2.2免疫信號在淋巴細胞間的轉導 在自身免疫疾病發生的過程中,淋巴細胞間的免疫信號轉導是一個重要環節。在這個過程中,抗原呈遞細胞將處理好的抗原提呈給T細胞,T細胞活化后刺激B細胞活化,B細胞活化后分化為漿細胞產生針對抗原的自身抗體,同時T細胞產生細胞因子調節抗體的產生。其中涉及許多與SLE易感性相關的基因。
2.2.1B細胞受體信號通路轉導
2.2.1.1具有錨蛋白重復序列1(BANK1)的B細胞支架蛋白 BANK1是編碼B細胞支架蛋白的一種基因。該基因編碼產物可以轉導B細胞信號,當B細胞抗原受體與抗原結合后,刺激BANK1與SRC家族B淋巴細胞激酶(BLK)基因結合,向胞內傳遞信號,并激活下游的基因。其多態性可能會導致B細胞耐受破壞或者產生持續的信號傳導。已有許多研究探討BANK1多態性與SLE的易感性,均顯示出二者之間極強的相關性,如rs10516487、rs71597109的多態性均與SLE存在較強的關聯[24-25]。
2.2.1.2蛋白酪氨酸磷酸酶非受體22型(PTPN22) PTPN22基因編碼的淋巴特異性酪氨酸激酶(LYP)的蛋白質,是T細胞活化的有力負向調節劑,它與C末端SRC酪氨酸激酶(CSK)相互作用,影響下游SRC家族酪氨酸激酶的活化狀態[26]。它的基因多態性與SLE、類風濕性關節炎、系統性硬化癥、自身免疫性甲狀腺疾病等多種自身免疫疾病密切相關,其中rs2476601是PTPN22中與疾病相關的主要SNP,與SLE有相關性[27]。
2.2.1.3CSK CSK參與SRC家族激酶的調節,主要在免疫細胞表面表達,其基因編碼的蛋白質與PTPN22結合后,在T、B淋巴細胞的活化中起重要作用。研究已確定CSK基因內含子的多態性與SLE的關聯性,其中rs34933034多態性增加了狼瘡的易感性,與SLE的發生有強關聯[26]。
2.2.1.4BLK SRC家族激酶是一類非受體酪氨酸激酶,到目前為止有9個成員,即SRC、YES、FYN、FGR、LCK、HCK、BLK、LYN和FRK,其中BLK與SLE的發生密切相關。
BLK原癌基因是SRC家族的酪氨酸激酶,參與細胞增殖和分化。BLK蛋白在B細胞受體信號傳導和B細胞發育中起作用。BLK基因存在多種基因多態性位點,其與SLE的易感性相關,其中包括rs13277113、rs2736340和rs2248932[28]。BLK多態性除了與SLE的發病相關以外,還有研究顯示其與SLE的疾病活動度相關。與其他基因型相比,rs13277113 G/A型和rs2736340 C/T型在活躍期SLE患者中顯著高于其他基因型[29]。
2.2.2T細胞受體信號通路轉導 T細胞接受抗原呈遞細胞提呈的抗原后,繼續提呈給B細胞,同時自身進行分化,其中輔助性T細胞(Th)2、Th17與SLE的發生密切相關。
2.2.2.1IL-17 研究顯示,IL-17與SLE的發生相關,Th17細胞產生細胞因子IL-17參與免疫調節,在許多自身免疫疾病中發揮重要作用[30]。目前有研究顯示,SLE患者體內IL-17的水平顯著高于健康對照,提示IL-17在SLE的發生過程中發揮作用[31]。除了水平升高之外,IL-17基因rs2275913的多態性也與SLE相關,并且影響疾病活動度[32]。
2.2.2.2IL-21 與IL-17類似,IL-21也是由Th17細胞分泌的細胞因子,參與免疫細胞增殖分化過程,在免疫應答中起重要作用,與多種自身免疫疾病相關。SLE患者體內IL-21水平及其mRNA表達量顯著升高,說明SLE的發生與IL-21在體內的過表達相關[33]。除此之外,IL-21的基因多態性也與SLE發病相關。已經有病例對照研究顯示,rs907715、rs2221903的多態性與SLE的發病相關[34]。SLE相關的常見基因及信號通路見表1。

表1 SLE相關的常見基因及信號通路
3 結 論
SLE目前是一種病因不清、發病機制復雜、臨床表現異質性大的自身免疫性疾病,發病過程涉及多個過程和通路。目前已明確的SLE致病相關基因有很多種,本文對其中較為常見的15個基因和2種表觀遺傳修飾方式進行了綜述(表1),涉及超過20種SNPs。除此之外,還有許多基因與SLE致病風險表現出一定的相關性,但尚不能明確,這些可能會對SLE致病有重要影響但未完全明確的基因值得進一步研究。隨著SLE遺傳學研究的深入,更多的SLE家系患者被發現,研究人員也更加重視SLE的家族聚集性,并且隨著分子診斷技術、遺傳學技術的進步,更多基因研究技術可供選擇,如檢測表達數量性狀位點(eQTLs)、RNA結合位點、蛋白質結合位點等,這為發現更多SLE易感基因提供了技術支持。
在不同地區及種族之間SLE易感基因不完全相同,疾病表現和嚴重程度也不完全一致,在對SLE易感基因進行研究時可以將種族或地區進行分層,從而探討不同人種的最強關聯基因或主要致病基因,再對重點基因進行功能學研究,這樣可以為SLE致病基因的研究和針對不同基因的靶向治療提供極大的幫助。此后,也可以分析SLE的疾病情況與不同基因之間的關系,研究疾病的早期產生、進展過程、活動度、并發癥及預后等不同過程中涉及的基因,進一步確定不同基因的主要影響方向,通過統計學方法和計算機技術建立數學模型,為以后臨床使用人工智能模型對SLE進行多基因分析及預測提供可能。
在之后的研究中,對已發現的易感基因分類研究,如按信號轉導通路、對疾病的影響方面等對各種基因進行歸類,再結合統計學分析方法,探索不同類別的相關基因與各臨床表現及亞型之間的關聯性。未來對SLE患者進行基因檢測,確定致病基因靶點,可以為SLE的精準治療提供實驗室依據,發展SLE治療的個體化,提高治療的有效性,減少治療不良反應,改善患者預后。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
