西花薊馬P450基因在繼代適應菜豆植株中的應答反應
2020-05-11 10:04:52郅軍銳岳文波
環境昆蟲學報 2020年2期
關鍵詞:差異
劉 利,郅軍銳,岳文波,謝 文,張 濤
(貴州大學昆蟲研究所,貴州省山地農業病蟲害重點實驗室,貴陽 550025)
寄主植物和植食性昆蟲在長期的進化過程中形成了一種相互對抗和彼此適應的機制。被昆蟲取食后,植物能產生防御反應抵御植食性昆蟲進一步取食為害,植食性昆蟲也能通過反防御機制適應寄主植物的防御,確保種群的生存和繁衍(彭露等,2010;劉勇等,2011;禹海鑫等,2015)。細胞色素P450是一類廣泛存在于生物體內的超家族蛋白,除與害蟲的抗藥性有關外(Schuler,2011;Mizutani and Masaharu,2012),在外源有毒物質如次生物質的解毒以及寄主植物的適應性中發揮著重要的作用(邱星輝,2014;陳澄宇等,2015;Liuetal.,2015a)。如煙粉虱Bemisiatabaci(謝文,2011)、二斑葉螨Tetranychusurticae(戴宇婷等,2012)、B型煙粉虱和Q型煙粉虱(楊金鍵等,2017)在取食不同寄主后,體內細胞色素P450活性發生改變迅速適應不同寄主。Peng等(2016)研究發現煙蚜Myzuspersicae的P450基因CYP6CY4 和CYP6CY3 的表達與次生物質煙堿密切相關。Zhu等(2016)發現馬鈴薯葉甲Leptinotarsadecemlineata取食植物毒素和藥劑處理的馬鈴薯后,P450s基因表達上調,參與植物毒素和藥劑的代謝。花椒毒素也可誘導美洲棉鈴蟲Helicoverpzea體內P450s基因CYP6B8 和CYP321A1以及珀鳳蝶PapiliopolyxenesCYP6B1基因的表達(Petersenetal.,2001;Lietal.,2004)。
西花薊馬Frankliniellaoccidentalis是世界性的重要入侵害蟲(Kirk and Terry,2003;Reitzetal.,2011),除直接取食和產卵危害,還能傳播多種植物病毒,造成巨大的經濟損失(謝永輝等,2013)。西花薊馬于2003年在北京首次報道其為害(張友軍等,2003),隨后在中國各地擴散(呂要斌等,2011),甚至西藏(王海鴻等,2013)、吉林(劉若思等,2015)和內蒙古中西部地區(高振江等,2017)也有其危害的報道,這可能與其對環境和寄主強大的適應能力有關。因此,探討西花薊馬對寄主植物的適應性,對揭示其入侵、傳播和擴散具有重要意義。
西花薊馬寄主植物廣泛,但對不同寄主的適應性不同。菜豆是西花薊馬的嗜食寄主,但菜豆不同部位對西花薊馬的生長發育和繁殖影響不同,西花薊馬對菜豆豆莢的嗜食性高于菜豆植株葉片(郅軍銳等,2010)。前人對西花薊馬適應寄主植物機制的報道主要集中在其取食不同寄主(Jensenetal.,2000;劉建業等,2017)或不同誘導的寄主植物后蟲體生理生化水平的變化(郅軍銳等,2016;蒲恒滸等,2018),而西花薊馬代謝相關基因(如P450s)的應答機制,及逐代適應的調控模式尚不明確。本文在前人研究的基礎上,通過實時熒光定量PCR分析了西花薊馬從菜豆豆莢轉換到菜豆植株適應不同世代后,其2齡若蟲和成蟲體內CYP4基因和CYP6相關基因表達量的變化,為深入探討西花薊馬對寄主植物適應機制提供依據。
1 材料與方法
1.1 材料
供試昆蟲:西花薊馬為人工氣候箱中長期繼代飼養(20代以上)的菜豆豆莢種群。條件為溫度25±1℃,相對濕度75%±5%,光周期14 h ∶10 h(L ∶D)。
供試植株:矮生菜豆Phaseolusvulgaris品種:金束鹿泰國地豆王(河北省辛集市盛農種子公司)。于人工氣候室內種植菜豆植株,條件為溫度25±1℃,相對濕度75%±5%,光周期14 h ∶10 h(L ∶D)。每營養缽(直徑10 cm,高9 cm)培育兩株清潔苗,待菜豆長至3~4真葉時,取長勢一致的菜豆植株作為供試寄主植物。生長期間不施用任何農藥。
1.2 方法
1.2.1菜豆植株上不同世代西花薊馬的飼養及2齡若蟲和成蟲的取樣
選取健康、長勢一致的菜豆植株置于人工氣候室的養蟲籠(86 cm×45 cm×41 cm)中,取菜豆豆莢上飼養的羽化后3 d并已交配的西花薊馬雌成蟲接到菜豆植株上,24 h后剔除所接所有成蟲,待西花薊馬產下的卵發育至2齡若蟲階段,每個養蟲籠取200頭若蟲為一個樣,放入1.5 mL離心管中,記為F1代2齡若蟲,并立即用液氮冷凍后保存于-80℃冰箱中備用,同時保留一定若蟲繼續在植株上飼養,至成蟲階段后取成蟲150頭放入1.5 mL離心管中,記為F1代成蟲。
取F1代已交配的雌成蟲,依照F1代相同的方法轉換到新的菜豆植株上,待發育到2齡若蟲和成蟲后進行取樣,分別記為F2代2齡若蟲和F2代成蟲。按照以上方法繼續取F3代2齡若蟲和F3代成蟲。每一個養蟲籠的薊馬為1個重復,共設置5個生物學重復。以取食豆莢的西花薊馬2齡若蟲和成蟲為對照,記為CK。
1.2.2西花薊馬總RNA的提取
參照Eastep? Super總RNA提取試劑盒說明書步驟提取西花薊馬2齡若蟲和成蟲的總RNA,并保存于-80℃冰箱內備用。
1.2.3cDNA第一條鏈的合成
參照RevertAid First Strand cDNA Synthesis Kit試劑盒說明書步驟合成cDNA第一條鏈,-20℃冰箱內保存備用。
1.2.4實時熒光定量(Realtime qPCR)分析
根據西花薊馬已克隆的5個CYP4基因(董紅剛等,2015)和EST數據庫中的4個CYP6基因CYP6-1(GQ290643)、CYP6-2(GQ29064)、CYP6-3(GT298397)和CYP6-4(GT298649)(謝志娟,2013)合成的qPCR特異性引物(見表1),以西花薊馬EF-1作為內參基因(Zhengetal.,2014)進行實時熒光定量PCR(RT-qPCR)。引物由生工生物工程(上海)股份有限公司合成。
反應在CFX96?實時熒光定量PCR儀(Bio-rad公司)上進行。反應體系10 μL:5.0 μL FastStart Essential DNA Green Master,正反向引物各0.5 μL,cDNA 1 μL,DEPC水3 μL。程序為:95℃預變性10 min,95℃變性30 s、60℃退火和延伸30 s,進行40個循環。每次生物重復進行3次技術重復。反應完成收集Ct值,分析溶解曲線。
1.3 數據分析
結果采用2-ΔΔCt法(Livak and Schmittgen,2001)進行基因相對表達量的計算。用Excel 2016和SPSS 19.0軟件對實驗數據進行統計和分析,不同世代間差異性分析用Duncan氏多重比較,2齡若蟲和成蟲間的差異采用獨立樣本t檢驗。

表1 RT-qPCR所用特異引物Table 1 Primers used in RT-qPCR
2 結果與分析
2.1 西花薊馬轉換到菜豆植株后體內CYP4基因差異表達分析
對于CYP4基因,西花薊馬從菜豆豆莢轉換到菜豆植株后,2齡若蟲體內CYP4-1和CYP4-2的表達量變化相似,均是在F1代顯著升高,分別為對照的4.37倍和2.31倍,而在F2和F3代其表達量均下降但仍高于對照,且F2和F3代間無顯著差異。2齡若蟲CYP4-3的表達量只有F1代顯著升高,其余世代與對照則無顯著差異。CYP4-4表達量僅在F3代顯著高于對照,為對照的13.61倍,其余世代與對照無顯著差異;CYP4-5表達量的變化呈現先下降后上升的變化,在F2代達到最低值,僅為對照的44.62%,且F1與F3代表達量相當,顯著低于對照,分別只有對照的73.04%和68.18%。
成蟲體內CYP4-1、CYP4-2和CYP4-3在F1代的表達量與對照相比均無顯著差異;而CYP4-1和CYP4-2的表達量在F2代顯著下降到最低,到F3代有所上升但仍顯著低于對照;CYP4-3的表達量則隨世代的增加逐漸降低;CYP4-4和CYP4-5的表達量在F1代顯著升高,而F2和F3代與F1代相比均顯著降低,且兩世代間差異不顯著,其中CYP4-4下降到對照的表達水平,而CYP4-5的表達量則顯著低于對照。
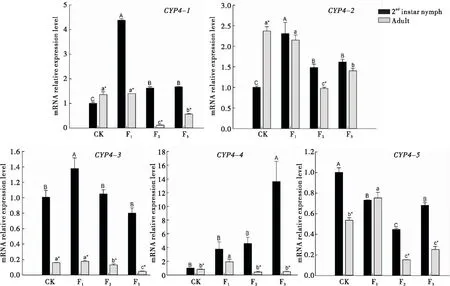
圖1 西花薊馬CYP4基因的相對表達量Fig.1 Relative expression levels of CYP4 gene in Frankliniella occidentalis注:圖中數據為平均值±標準誤,大寫字母表示西花薊馬2齡若蟲體內CYP4基因表達量在不同世代間顯著差異;小寫字母表示西花薊馬成蟲體內CYP4基因表達量在不同世代間的差異顯著性(Duncan氏多重比較,P<0.05)。*表示同一世代西花薊馬體內CYP4基因的表達量在2齡若蟲和成蟲之間存在顯著差異(t 檢驗,P<0.05)。下圖同。Note:Data in the figure are expressed as mean±SE.Capital letters indicate significant differences at the level of 0.05 of gene expression in 2nd instar nymphs among different generations,while lowercase letters indicate significant differences at the level of 0.05 of gene expression in adults among different generations (Duncan’s multiple range test).* indicated that there were significant differences at the level of 0.05 between the 2nd instar nymphs and adults at the same generation (t test).The same for the following figures.
同一個基因的表達量在西花薊馬成蟲與2齡若蟲間不同。取食菜豆豆莢的西花薊馬成蟲體內CYP4-1和CYP4-2的表達量顯著高于2齡若蟲,而CYP4-3、CYP4-4和CYP4-5的表達量相反,均顯著低于2齡若蟲的。轉換到菜豆植株后只有CYP4-2在F1和F3代,以及CYP4-4和CYP4-5在F1代的西花薊馬2齡若蟲和成蟲之間無顯著差異,其余CYP4基因在2齡若蟲的表達量均顯著高于成蟲。
2.2 西花薊馬轉換到菜豆植株后體內CYP6基因差異表達分析
從豆莢轉換到菜豆植株后,西花薊馬2齡若蟲體內CYP6基因的表達量在不同世代間的變化不同。CYP6-1的表達量在取食菜豆植株后只有F2代顯著降低,而CYP6-2表達量只有在F3代才顯著升高;CYP6-3 的表達量在取食菜豆植株后均顯著低于對照,且在F2代最低;而CYP6-4的表達量只有在F1代才明顯升高。
轉換到菜豆植株后,西花薊馬成蟲體內CYP6-1、CYP6-2和CYP6-3的表達量變化相似,都是在F1代顯著下降,與對照相比分別降低了39.55%、39.54%和26.90%,且3個基因在F2和F3代間的表達量均無明顯差異,但均顯著低于F1代,CYP6-4的表達量在F1代顯著升高,但F2、F3代與對照無顯著差異。
西花薊馬從菜豆豆莢轉換到菜豆植株后,除CYP6-3的表達量在F1代的2齡若蟲和成蟲間差異不顯著外,在其余情況下均是2齡若蟲體內的表達量顯著高于成蟲。
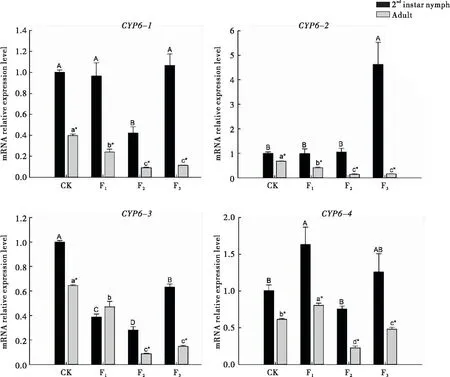
圖2 西花薊馬CYP6基因的相對表達量Fig.2 Relative expressionlevels of CYP6 gene in Frankliniella occidentalis
3 結論與討論
細胞色素P450是一類由CYP基因編碼的多基因超家族酶(Werckreichhart and Feyereisen,2000;Feyereisen,2006),對昆蟲體內多種內源性和外源性物質的氧化代謝起著重要作用,涉及昆蟲的生長發育、營養、對植物毒素的耐受性和對殺蟲劑的抗藥性等(Scott and Wen,2001;Bonoetal.,2008)。本研究發現西花薊馬從嗜食性很高的菜豆豆莢轉換到嗜食性次之的菜豆植株后,2齡若蟲體內CYP4-1、CYP4-2、CYP4-3、CYP6-4和成蟲體內CYP4-4、CYP4-5、CYP6-4的表達量均在F1代顯著上升,以迅速適應新的寄主。這可能是由于西花薊馬轉換到菜豆植株后,誘導植物產生有毒次生物質和防御蛋白,對西花薊馬產生不利影響(田甜等,2014;郅軍銳等,2016;岳文波等,2018),從而導致其解毒酶基因發生不同的變化。前人也有類似的結果,如周新改(2011)發現煙粉虱轉換到新寄主甘藍后,蟲體P450基因表達量上升。槲皮素、呋喃香豆素和花椒毒素等多種植物次生物質也能夠誘導昆蟲P450不同基因的表達(Lietal.,2004;Maoetal.,2009;陳澄宇等,2015)。彭天飛(2016)發現CYP6CY3 參與代謝次生物質煙堿,其表達量與煙蚜寄主適應性相關。棉鈴蟲Helicoverpaarmigera在次生物質槲皮素誘導后,CYP337B1 和CYP6B6 表達量顯著升高(Liuetal.,2015b)。Yang等(2011)研究發現褐飛虱Nilaparvatalugens取食抗性品種水稻后,CYP6家族基因CYP6CS1和CYP6CW1 被顯著誘導。
西花薊馬2齡若蟲體內CYP4-1、CYP4-2、CYP4-3、CYP6-4和成蟲體內CYP4-4、CYP4-5、CYP6-1、CYP6-2、CYP6-3的表達量均在F2代之后達到穩定,這與煙粉虱寄主轉換后體內α-NA羧酸酯酶和谷胱甘肽S-轉移酶活性在F2代基本穩定的結果相呼應(周福才等,2010),這可能是由于昆蟲轉換到新的寄主或食物后,逐漸適應新的取食環境,致使生理調控達到平衡。而桃蚜取食蠶豆后CYP6a13和CYP6a14的表達量平衡點延遲,分別在第6和7代后才達到穩定(朱經云,2017)。P450表達量變化的差異可能與不同基因、昆蟲種類和寄主種類有關。CYP4-5、CYP6-1、CYP6-3在2齡若蟲期的表達量和CYP4-1、CYP4-2、CYP6-4在成蟲期的表達量變化相似,除CYP6-1在F3代恢復到對照水平外,其他基因表達量在F3代時均低于對照,這說明昆蟲解毒酶基因在適應取食新的寄主后,不同基因的表達存在特異性,彼此協調以迅速適應新的環境。
本研究結果還顯示,西花薊馬轉換到菜豆植株除后CYP4-2的表達量在F1和F3代,以及CYP4-4、CYP4-5和CYP6-3的表達量在F1代兩個蟲態間差異不顯著外,其余情況下均是2齡若蟲體內的表達量顯著高于成蟲的,這可能由于2齡若蟲取食量較成蟲大,或是由于成蟲需要節約能量進行后代繁殖所致。前人的研究也表明P450基因在同一昆蟲不同蟲態體內的表達量不同,周興隆等(2016)研究發現,二斑葉螨抗性品系中CYP392E10和CYP392A6的表達量在若螨階段上調,成螨則不顯著,而CYP392A16的表達量各個發育階段都上調,CYP392D8則在除若螨外的其他階段都上調。煙粉虱噻蟲嗪抗性品系中P450基因p_06013和p_00988在4齡若蟲過量表達,而p_05916和p_00478表達量很低且在成蟲期過量表達,p_00059和p_00428也在雌成蟲階段過量表達(楊妮娜等,2016)。可見環境脅迫能明顯影響昆蟲不同發育階段P450基因的表達量,并且因基因、蟲態和環境而異。
本研究探討了P450基因在西花薊馬豆莢種群轉換到菜豆植株后逐代適應中的作用,為進一步研究西花薊馬的適應性提供了依據。但寄主植物和植食性昆蟲的相互作用十分復雜,不同食物的“口感”、營養成分以及植物的防御反應(植物次生物質的種類和濃度等)等都會對昆蟲造成不同影響,昆蟲可通過取食行為、生長發育、生理生化及分子防御等以適應不同寄主(彭露等,2015)。因此西花薊馬對寄主的適應機制還有待進一步深入探討。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
