二氧化碳聯(lián)合核桃殼提取液促進(jìn)單針藻Monoraphidium sp.QLZ-3的生長(zhǎng)和油脂積累
2020-05-08 10:24:40邢海亮董訓(xùn)贊韓本勇耿樹(shù)香寧德魯馬婷余旭亞
化工進(jìn)展 2020年4期
關(guān)鍵詞:油脂
邢海亮,董訓(xùn)贊,3,韓本勇,耿樹(shù)香,寧德魯,馬婷,余旭亞
(1昆明理工大學(xué)生命科學(xué)與技術(shù)學(xué)院,云南昆明650500;2云南省林業(yè)與草原科學(xué)院,云南昆明650051;3云南貴澳農(nóng)業(yè)科技集團(tuán),云南昆明650100)
近年來(lái),由于微藻具有光合效率高、生長(zhǎng)周期短、油脂含量高等優(yōu)勢(shì),被公認(rèn)為制備生物柴油的新型生物質(zhì)能源[1-2]。然而,較高的生產(chǎn)成本及較低的生物量產(chǎn)率和油脂產(chǎn)率限制了利用微藻制備生物柴油的工業(yè)化發(fā)展[3-4]。利用廢棄物中的營(yíng)養(yǎng)物質(zhì)培養(yǎng)微藻,可以有效地降低微藻生產(chǎn)成本,同時(shí)廢棄物得到資源化利用,成為研究熱點(diǎn)[5]。
核桃是營(yíng)養(yǎng)豐富的堅(jiān)果,種植廣泛,核桃殼作為核桃產(chǎn)品加工過(guò)程中的副產(chǎn)物,可以用于色素的提取、制作活性炭和拋光材料等[6-7]。然而大部分核桃殼卻被就地焚燒,不僅造成了資源的浪費(fèi),還會(huì)帶來(lái)嚴(yán)重的環(huán)境污染。研究指出,核桃殼中含有多種微量元素及豐富的多酚物質(zhì)[8]。多酚作為一種抗氧化劑,具有較強(qiáng)的抗氧化性[9]。Zhao 等[10]研究指出,抗氧化劑可以通過(guò)調(diào)節(jié)與油脂合成相關(guān)的酶的活性,進(jìn)而有效促進(jìn)非生物脅迫下微藻中油脂的積累。此外,在微藻的培養(yǎng)過(guò)程中,通入適當(dāng)濃度的CO2,有利于提高微藻的光合作用效率,促進(jìn)微藻生物量的積累[11]。有研究指出,在BG-11無(wú)碳培養(yǎng)基中,當(dāng)外源通入5% 的CO2時(shí),小球藻FACHB-1580和柵藻FACHB-1618最大生物量達(dá)到3.5g/L 和5.4g/L,分別是對(duì)照組的1.41 倍和1.46倍[12]。然而,核桃殼提取液(walnut shell extracts,WSE)聯(lián)合CO2用于微藻培養(yǎng)的研究鮮有報(bào)道,研究CO2對(duì)WSE中微藻生長(zhǎng)和油脂積累的影響,有利于了解CO2作用下WSE中微藻生長(zhǎng)和油脂合成的相關(guān)機(jī)制。
本文以單針藻Monoraphidium sp. QLZ-3 為對(duì)象,以WSE為培養(yǎng)基,研究了外源通入CO2的條件下,微藻生物量產(chǎn)率和油脂產(chǎn)率的變化及對(duì)WSE 中多酚的吸收。同時(shí)對(duì)微藻細(xì)胞內(nèi)核酮糖1,5-二磷酸羧化酶基因(ribulose 1,5-bisphosphate carboxylase/oxygenase,rbcL)相對(duì)表達(dá)水平和與油脂合成相關(guān)酶的活性進(jìn)行了檢測(cè)。將微藻培養(yǎng)與核桃殼的利用相結(jié)合,為提高微藻的生物量產(chǎn)率和油脂產(chǎn)率,降低微藻培養(yǎng)成本,以及核桃殼的環(huán)境友好的資源化利用提供一定的指導(dǎo)依據(jù)。
1 材料與方法
1.1 原料與培養(yǎng)基的制備
以單針藻Monoraphidium sp. QLZ-3 為研究對(duì)象(本實(shí)驗(yàn)室篩選、保存)[13]。稱(chēng)取適量的核桃殼(云南省林業(yè)科學(xué)院提供)與10倍質(zhì)量的蒸餾水混合,在95℃條件下提取4h,得到褐色的WSE,經(jīng)8層紗布過(guò)濾后,添加NaNO3(1.16g/L),調(diào)整pH至6.8~7.0,分裝至生物反應(yīng)器(?0.06m×h0.51m)中,121℃滅菌20min。將單針藻Monoraphidium sp.QLZ-3接入上述培養(yǎng)基中(初始接種量為0.4g/L),分別通入不同濃度CO2(空氣、4%、8%、12%和16%),氣體流速為0.5L/min,光照強(qiáng)度6500lux,培養(yǎng)溫度25℃±1℃,每組設(shè)置3個(gè)平行。
1.2 儀器與設(shè)備
FA2004N 分析天平,LDZX-50KBS 滅菌鍋,VS-840-1超凈工作臺(tái),Ultrospec 2100pro紫外可見(jiàn)分光光度計(jì),RF-540 熒光分光光度計(jì)(Shimadzu),5804R 高速低溫離心機(jī)(德國(guó)Eppendorf 公司),1730R 高速冷凍離心機(jī)(丹麥Labogene Scanspeed公司),F(xiàn)D5-12冷凍干燥機(jī)(西盟國(guó)際集團(tuán)),Agilent 7890A系列氣相色譜儀(Agilent Technologies)。
1.3 生物量、油脂含量的測(cè)定[14]
(1)生物量 每天取10mL(V)藻液,3500r/min 離心10min,去上清液,冷凍干燥后,稱(chēng)取藻粉質(zhì)量(w1)。微藻生物量Y(g/L)的計(jì)算如式(1)。
Y = w1/V (1)
(2)油脂含量 將新鮮的藻液冷凍干燥后,稱(chēng)取0.3~0.5g(m1)干藻粉與2 倍質(zhì)量的石英砂混合,充分研磨40min,用3mL 氯仿甲醇溶液(2∶1,體積比)提取油脂,于25℃、150r/min 條件下振蕩提取30min后,離心(5000r/min,10min),收集上清液。重復(fù)上述提取操作3 次,合并上清液,置于預(yù)先干燥稱(chēng)重的離心管(w2)中,40℃干燥至恒重(w3),油脂含量η(%)計(jì)算如式(2)。

1.4 CO2固定效率及氣液傳質(zhì)系數(shù)的測(cè)定
二氧化碳固定效率依據(jù)式(3)計(jì)算[15]。

式中,F(xiàn)CO2為CO2固定效率,mg·L-1·d-1;Pb為生物量產(chǎn)率,mg·L-1·d-1;a 為單位生物量固定CO2的質(zhì)量。
氣液傳質(zhì)系數(shù)[16]測(cè)定方法為:先將光生物反應(yīng)器中不含微藻的培養(yǎng)基以99.99%的氮?dú)馄貧?.5h后,再以不同濃度的CO2曝氣,培養(yǎng)基溫度為25±1℃,測(cè)量60min 內(nèi)培養(yǎng)基中溶解CO2濃度的變化,每5min記錄1次,通過(guò)氣液傳質(zhì)方程和亨利定律計(jì)算傳質(zhì)系數(shù)KLa。計(jì)算過(guò)程如式(4)。
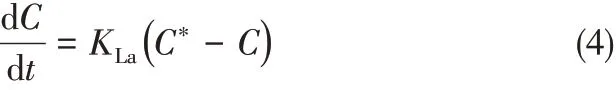
1.5 rbcL基因表達(dá)分析
使用Primer5.0 設(shè)計(jì)引物,由上海生工生物工程技術(shù)服務(wù)有限公司合成18S和rbcL酶基因的上下游擴(kuò)增引物(表1),并以此進(jìn)行熒光定量PCR。
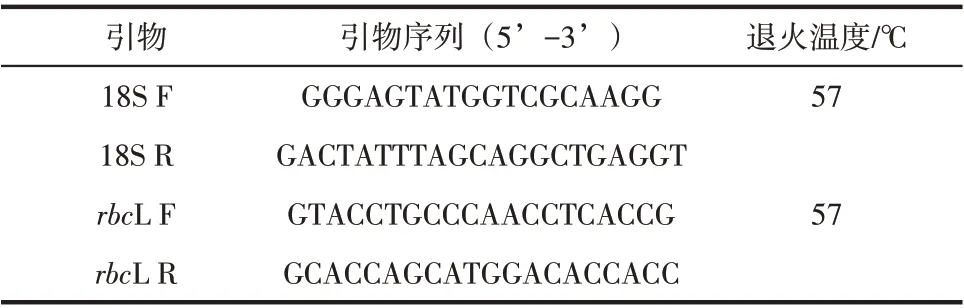
表1 酶基因熒光定量引物
Trizol 法提取不同濃度CO2處理下單針藻Monoraphidium sp. QLZ-3 內(nèi)的RNA,利用逆轉(zhuǎn)錄試劑盒(TaKaRa)將RNA 逆轉(zhuǎn)錄合成cDNA,以其為模板進(jìn)行RT-PCR 擴(kuò)增,檢測(cè)不同濃度CO2處理下單針藻rbcL 基因表達(dá)的變化。通過(guò)ABI 7500熒光定量?jī)x對(duì)rbcL 基因的表達(dá)進(jìn)行定量,RT-PCR的數(shù)據(jù)結(jié)果用2-ΔΔCT(Livak)的方法處理分析。以18s基因作為內(nèi)標(biāo)以調(diào)節(jié)RNA的用量和循環(huán)數(shù),使內(nèi)標(biāo)基因在誘導(dǎo)條件下的表達(dá)豐度一致,最終得到基因表達(dá)量之間的倍數(shù)關(guān)系。
1.6 多酚含量測(cè)定[17]
WSE 中總多酚含量采用Folin-Ciocalteu 法測(cè)定:稱(chēng)取干燥至恒重的沒(méi)食子酸10.0mg,加入到蒸餾水中定容至100mL。分別取0.1mL、0.2mL、0.3mL、0.4mL、0.5mL、0.6mL、0.7mL、0.8mL 和0.9mL 沒(méi)食子酸溶液于10mL 的比色管中,加入Folin-Ciocalteu 試劑0.5mL,1min 后加入碳酸鈉溶液1.5mL(200g/L),用蒸餾水定容至刻度線處,混勻,40℃保溫2h 后,迅速冷卻。在760nm 處測(cè)定吸光度,建立標(biāo)準(zhǔn)曲線。按照上述方法測(cè)定樣品中多酚的含量。
1.7 酶活性測(cè)定[18]
單針藻Monoraphidium sp. QLZ-3 中蘋(píng)果酸酶(malic enzyme,ME)、乙酰輔酶A 羧化酶(acetyl coenzyme A carboxylase,ACCase)和磷酸烯醇式丙酮 酸 羧 化 酶 (phosphoenolpyruvatecarboxylase,PEPC)的活性使用比色定量試劑盒(蘇州科銘生物技術(shù)有限公司)測(cè)定。
1.8 脂肪酸組成分析
將2mL 3%的硫酸-甲醇(體積比)溶液加入到烘干的油脂中,70℃回流2h 后,加入2mL 正己烷震蕩提取4h,取正己烷相進(jìn)行氣相色譜-質(zhì)譜(GC-MS)分析[19]。
1.9 統(tǒng)計(jì)分析
本文全部實(shí)驗(yàn)均設(shè)置3 組平行,利用ANOVA(SPSS19.0)一步法分析實(shí)驗(yàn)數(shù)據(jù),P<0.05表示差異顯著,后文中用“*”表示;P<0.01表示差異極顯著,后文中用“**”表示。
2 實(shí)驗(yàn)結(jié)果與討論
2.1 CO2對(duì)WSE中單針藻生物量的影響
不 同 濃 度 的 CO2對(duì) WSE 中 單 針 藻Monoraphidium sp. QLZ-3 生長(zhǎng)的影響如圖1 所示。在培養(yǎng)過(guò)程中,隨著CO2濃度的不斷提高,微藻的生物量逐漸增加,當(dāng)CO2濃度達(dá)到12%時(shí),微藻的生物量達(dá)到最高,為1.18g/L,是對(duì)照組的1.33倍。繼續(xù)增加CO2的濃度,微藻的生物量出現(xiàn)下降趨勢(shì)。近期研究指出,向糖蜜酒精廢醪液中添加褪黑素,單針藻Monoraphidium sp. QLY-1 的生物量達(dá)到1.22g/L[1],相比對(duì)照組提高了41.82%;在80mg/L黃腐酸的作用下,微藻的生物量提高了52.26%[9]。諸多研究表明,在微藻的生長(zhǎng)過(guò)程中,通入適當(dāng)濃度的CO2,可以促進(jìn)微藻光合作用的進(jìn)行,使得藻細(xì)胞快速生長(zhǎng)[11,20]。有研究指出,當(dāng)向生活污水中通入2.5%的CO2時(shí),柵藻的生物量達(dá)到1.37g/L,是對(duì)照組的1.54 倍[21]。適宜濃度的CO2有助于微藻的生長(zhǎng),但當(dāng)CO2的濃度超過(guò)一定的范圍時(shí),微藻的生長(zhǎng)就會(huì)受到抑制[22]。這與本實(shí)驗(yàn)結(jié)果一致,將WSE中的CO2濃度增加至16%時(shí),微藻的生長(zhǎng)受到抑制(圖1)。這是由于持續(xù)通入高濃度的CO2會(huì)迅速降低溶液pH,可能抑制相關(guān)酶活性,從而使微藻的生物量降低[12]。
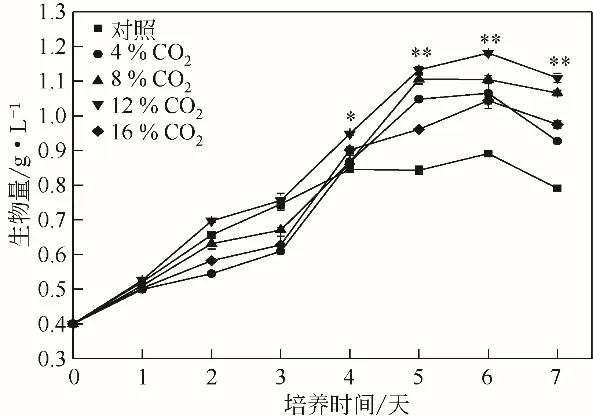
圖1 不同濃度CO2對(duì)WSE中單針藻Monoraphidium sp.QLZ-3生物量的影響
2.2 CO2 對(duì)WSE 中單針藻油脂及生物量產(chǎn)率的影響
如圖2 所示,當(dāng)WSE 中CO2的濃度為12%時(shí),單針藻Monoraphidium sp.QLZ-3 的油脂含量最高,為49.54%,是對(duì)照組的1.18 倍。此時(shí),生物量產(chǎn)率和油脂產(chǎn)率為196.85mg/(L·d)和97.52mg/(L·d),分別是對(duì)照組的1.33 倍和1.57 倍。相關(guān)研究指出,利用養(yǎng)豬廠廢水培養(yǎng)微藻,油脂含量最高可達(dá)28.8%,相比對(duì)照組提高了53.76%[23];利用水產(chǎn)養(yǎng)殖廢水培養(yǎng)柵藻LX1,其油脂含量可達(dá)到31.6%[24]。相關(guān)研究表明,由于微藻中誘導(dǎo)轉(zhuǎn)運(yùn)機(jī)制的存在,適當(dāng)濃度的CO2可以促進(jìn)微藻的生長(zhǎng)和積累脂類(lèi)[25]。當(dāng)BG-11 培養(yǎng)基中通入5% 的CO2時(shí),柵藻Scenedesmus sp. FACHB-1600 的油脂產(chǎn)率達(dá)到104.163mg/(L·d),相比對(duì)照組提高了51.15%[22]。本實(shí)驗(yàn)結(jié)果表明,適宜濃度的CO2有助于WSE中單針藻Monoraphidium sp.QLZ-3的生長(zhǎng)油脂積累,提高生物量產(chǎn)率和油脂產(chǎn)率。
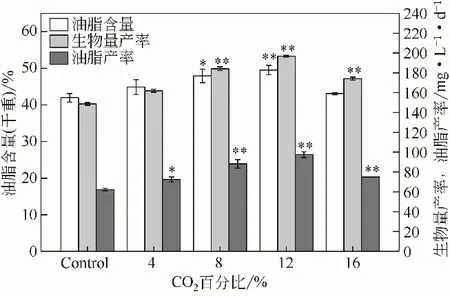
圖2 不同濃度CO2對(duì)微藻油脂含量、生物量產(chǎn)率和油脂產(chǎn)率的影響
此外,WSE的制備成本為CNY25.05/t(天然氣價(jià)格為CNY2.4/m3,水價(jià)格為CNY3.45/t),在12%的CO2作用下,微藻生物量和油脂的原料成本分別為CNY21.23/kg 和CNY42.85/kg。研究指出,利用傳統(tǒng)培養(yǎng)基BG-11 培養(yǎng)柵藻Scenedesmus SDEC-13時(shí),微藻生物量和油脂培養(yǎng)的原料成本則分別為CNY144.00/kg和CNY709.01/kg[26]。結(jié)果表明,WSE聯(lián)合CO2用于微藻的培養(yǎng)可以降低微藻生物柴油制備的原料成本。
2.3 CO2固定效率及傳質(zhì)效率
不同濃度的CO2對(duì)WSE 中傳質(zhì)系數(shù)(KLa)及微藻對(duì)CO2的固定效率的影響如圖3 所示。在微藻的培養(yǎng)過(guò)程中,隨著CO2濃度的增加,KLa值逐漸增加。當(dāng)CO2的濃度為12%時(shí),KLa值為0.073min-1,此時(shí)CO2的固定效率最高,達(dá)到0.36g CO2/(L·d)。結(jié)果表明,當(dāng)CO2的固定效率最高時(shí),單針藻Monoraphidium sp.QLZ-3的生物量產(chǎn)率及油脂產(chǎn)率達(dá)到最大(圖2)。而當(dāng)CO2的濃度為16%時(shí),雖然KLa值增加至0.081min-1,但CO2的固定效率下降至0.36g CO2/(L·d)(圖3),微藻的生物量產(chǎn)率和油脂產(chǎn)率也分別下降至174.05mg/(L·d)和74.97mg/(L·d)(圖2),這可能是由于較高的CO2濃度限制了微藻的生長(zhǎng)[27]。本實(shí)驗(yàn)結(jié)果顯示,當(dāng)KLa值為0.073min-1時(shí),WSE 中有足夠的持續(xù)的CO2供給微藻的生長(zhǎng),而當(dāng)KLa值增加0.081min-1時(shí),由于CO2濃度過(guò)高而降低了其固定效率,從而使微藻的生長(zhǎng)(圖1)和油脂積累(圖2)降低。
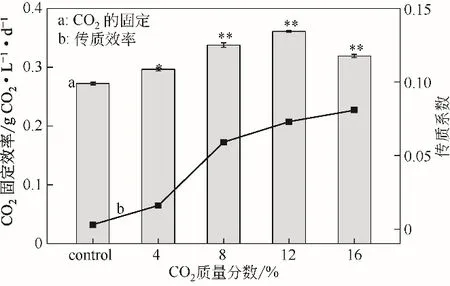
圖3 不同濃度CO2在WSE中對(duì)CO2的固定(柱形圖)及傳質(zhì)效率(折線圖)的影響
2.4 WSE中多酚含量變化
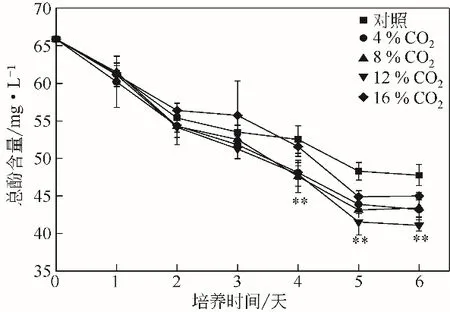
圖4 不同濃度CO2下WSE中多酚含量變化
多酚具有很強(qiáng)的抗氧化特性,可以減緩細(xì)胞的氧化損傷[9]。Zhao 等[10]研究指出,抗氧化劑可以通過(guò)調(diào)節(jié)油脂合成相關(guān)途徑,進(jìn)而促進(jìn)微藻中油脂的積累。由圖4可知,在微藻的培養(yǎng)過(guò)程中,WSE中的多酚含量逐漸降低。Lu等[28]指出,多酚是一種微量營(yíng)養(yǎng)元素,可被細(xì)胞吸收利用。在CO2的濃度為12%的條件下,WSE 中的多酚含量從65.87mg/L 下降至41.09mg/L,相比對(duì)照組降低了16.00%。圖1表明,12%的CO2促進(jìn)了微藻的生長(zhǎng),WSE中更多的多酚被單針藻Monoraphidium sp.QLZ-3吸收(圖4),從而促進(jìn)了微藻中油脂含量的積累(圖2)。Paz-Yépez 等[17]研究指出,多酚可以阻礙自由基與脂質(zhì)分子的結(jié)合,從而減少由于脂質(zhì)氧化引起的細(xì)胞損傷,促進(jìn)油脂的積累。褪黑素作為一種抗氧化劑(melatonin,MT),1μmol/L 的MT作用下,單針藻Monoraphidium sp. QLY-1 的油脂含量達(dá)到49.6%,是同等條件下對(duì)照組的1.32倍[29]。Che等[18]研究表明,抗氧化劑促進(jìn)微藻的生長(zhǎng)和油脂的積累,與油脂合成相關(guān)酶的活性及酶基因的表達(dá)有關(guān)。
2.5 CO2 對(duì)單針藻中與油脂合成相關(guān)酶活性的影響
ACCase 是一種依賴(lài)生物素的變構(gòu)羧化酶,主要催化依賴(lài)ATP 的乙酰輔酶A(acetyl coenzyme A)轉(zhuǎn)化為丙二酰輔酶A,是脂肪酸合成過(guò)程中的限速酶[1,28]。在CO2的作用下,單針藻Monoraphidium sp.QLZ-3 中,ACCase 的活性變化如圖5 所示,隨著CO2濃度的增加,藻細(xì)胞內(nèi)ACCase 的活性逐漸增強(qiáng),當(dāng)CO2濃度的增加至12%時(shí),ACCase 的活性最大,達(dá)到對(duì)照組的1.23~1.38 倍。繼續(xù)增加CO2的濃度,ACCase的活性開(kāi)始下降。前人研究指出,由ACCase 催化生成的丙二酰輔酶A 可以有效抑制脂肪酸的氧化[30]。通過(guò)上調(diào)ACCase 的活性,可以促進(jìn)衣藻Chlamydomonas reinhardtii 油脂含量的提高[1]。本實(shí)驗(yàn)研究結(jié)果顯示,向WSE中通入適宜濃度的CO2,可以提高單針藻Monoraphidium sp.QLZ-3中ACCase的活性,促進(jìn)油脂的積累。
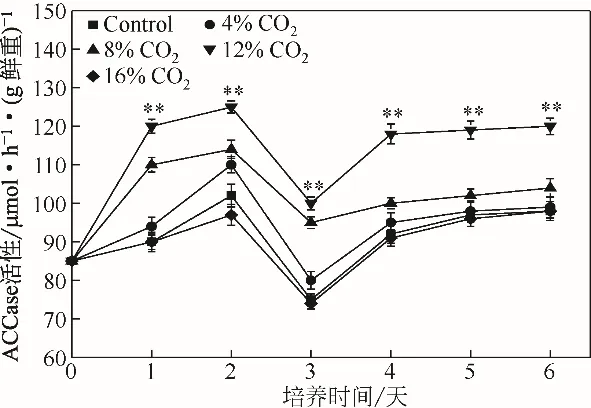
圖5 不同濃度CO2對(duì)微藻中ACCase活性的影響
蘋(píng)果酸在ME 的作用下降解為丙酮酸,還原NADP+,為脂肪酸的合成提供足夠的NADPH[1]。如圖6所示,CO2促進(jìn)了微藻中ME活性的提高。培養(yǎng)第2 天時(shí),ME 活性的提高最大,是對(duì)照組的2.60倍。在脂肪酸合成的過(guò)程中,脂肪酸合成酶可利用由ME 還 原 得 到 的NADPH[31]。 在 單 針 藻Monoraphidium sp. QLZ-3 的培養(yǎng)過(guò)程中,ME 的活性始終高于對(duì)照組(圖6),這就為微藻油脂的合成提供了足夠的NADPH,促進(jìn)了微藻油脂的積累(圖2)。Xue 等[32]研究指出,通過(guò)提高M(jìn)E 的活性,三角褐指藻的油脂含量提高了2倍。這與本實(shí)驗(yàn)結(jié)果一致,CO2促進(jìn)了微藻中ME 的活性,進(jìn)而促進(jìn)了微藻油脂含量的提高。
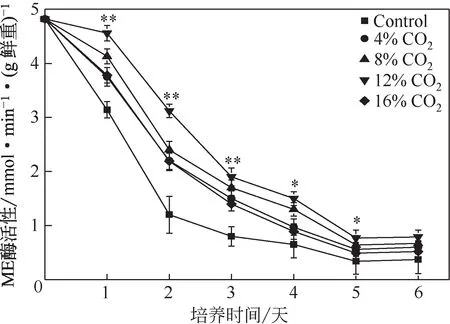
圖6 CO2作用下微藻中ME活性的變化
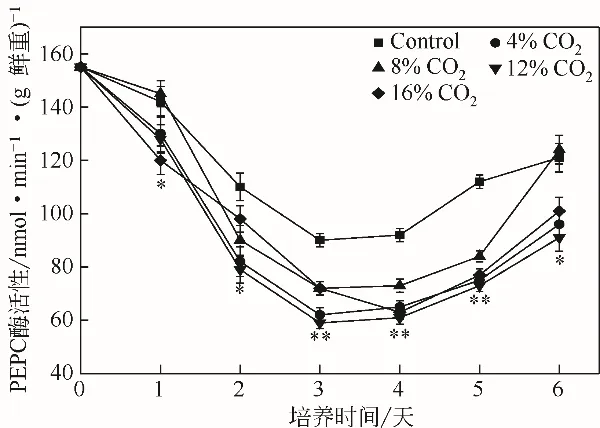
圖7 不同濃度CO2對(duì)單針藻Monoraphidium sp.QLZ-3中PEPC活性的影響
PEPC 是光合作用C4途徑中的關(guān)鍵酶之一,催化磷酸烯醇式丙酮酸的β-C羧基化并生成草酰乙酸和無(wú)機(jī)磷酸,它與ACCase 之間存在著底物的競(jìng)爭(zhēng)[33-34]。在微藻培養(yǎng)過(guò)程中,實(shí)驗(yàn)組和對(duì)照組中PEPC 的活性均出現(xiàn)先降低后增加的趨勢(shì),但實(shí)驗(yàn)組中的PEPC活性始終低于對(duì)照組(圖7)。培養(yǎng)前期,ACCase和ME的活性較高(圖5和圖6),油脂合成較快,PEPC的活性逐漸下降(圖7),培養(yǎng)后期,ACCase和ME的活性較低(圖5和圖6),油脂緩慢合成,PEPC活性開(kāi)始逐漸增高。Ghosh等[25]研究指出,通過(guò)上調(diào)衣藻Chlamydomonas reinhardtii中PEPC的活性,其油脂含量下降了37%。而PEPC基因的敲除,促進(jìn)了三角褐指藻Phaeodactylum tricornutum中油脂的積累[35]。本實(shí)驗(yàn)結(jié)果表明,CO2可以降低單針藻Monoraphidium sp. QLZ-3 中PEPC的活性,促進(jìn)微藻油脂的積累。
2.6 CO2對(duì)WSE中單針藻rbcL基因表達(dá)量的影響
單針藻Monoraphidium sp. QLZ-3 中rbcL 基因相對(duì)表達(dá)量的變化見(jiàn)圖8。在CO2的作用下,微藻細(xì)胞內(nèi)rbcL 基因的相對(duì)表達(dá)量均有不同程度的增加。與對(duì)照組相比,當(dāng)CO2的濃度為12%時(shí),在培養(yǎng)的1~3 天,rbcL 基因的相對(duì)表達(dá)量從1.64倍增加至2.63倍,隨后開(kāi)始下降,到第6天時(shí)下降至1.49 倍。Ikaran 等[36]研究表明,rbcL 基因與CO2的固定有關(guān),其表達(dá)量的上調(diào)可以加快光合作用固碳。圖8 表明,12%的CO2作用下微藻中rbcL 基因表達(dá)量上調(diào),促進(jìn)了微藻對(duì)CO2的吸收,為微藻的生長(zhǎng)提供更多的碳骨架,從而促進(jìn)了微藻的生長(zhǎng)(圖1)和CO2的固定(圖3)。Che 等[18]研究表明,微藻固定的CO2用于合成糖、脂質(zhì)等生物大分子。缺氮條件可以促進(jìn)小球藻chlorella vulgaris var L3中rbcL 基因表達(dá)量的上調(diào),油脂含量增加了25%[36]。本實(shí)驗(yàn)結(jié)果顯示,12%的CO2上調(diào)了rbcL基因表達(dá)量,促進(jìn)了CO2的固定、微藻的生長(zhǎng)及油脂的積累。
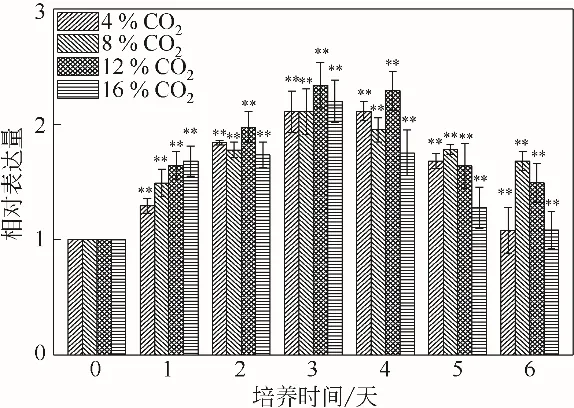
圖8 不同濃度CO2對(duì)單針藻Monoraphidium sp.QLZ-3的rbcL基因相對(duì)表達(dá)量影響
2.7 WSE中單針藻脂肪酸的組成分析
單針藻Monoraphidium sp.QLZ-3脂肪酸的組成如表2 所示,C16∶0(16 表示脂肪酸碳鏈的長(zhǎng)度,0表示碳鏈上不飽和鍵的個(gè)數(shù))、C18∶1、C18∶2和C18∶3是微藻脂肪酸的主要成分。與對(duì)照組相比,12%的CO2作用下,微藻中的C16∶0 含量增加了36.27%,而C18∶2的含量則減少了34.15%,C18∶1和C18∶3 的含量基本保持不變,這就導(dǎo)致了實(shí)驗(yàn)組中飽和脂肪酸(saturated fatty acid,SFAs)含量上升,多不飽和脂肪酸(polyunsaturated fatty acid,PUFA)含量下降,單不飽和脂肪酸(monounsaturated fatty acid,MUFA)含量保持不變。12%的CO2作用下的實(shí)驗(yàn)組中不飽和度(degree of unsaturation,DU)為103.83,相比對(duì)照組下降了15.48%,可作為生產(chǎn)生物柴油的原料[37]。此外,WSE中所培養(yǎng)微藻脂肪酸的長(zhǎng)鏈飽和因子(long chain saturation factor,LCSF)和冷濾點(diǎn)(cold filter plugging point,CFPP)如表2 所示。LCSF 是用來(lái)計(jì)算生物柴油的CFPP值的,CFPP值越低,生物柴油的低溫流動(dòng)性能越好。由于各國(guó)所處的環(huán)境不同,對(duì)CFPP 值也有不同的規(guī)定,而歐洲柴油標(biāo)準(zhǔn)并未對(duì)生物柴油的CFPP 值提出具體的要求,但我國(guó)(GB/T25199—2010)規(guī)定其值不高于-5~12℃。本實(shí)驗(yàn)結(jié)果表明,以WSE 為培養(yǎng)基,培養(yǎng)微藻得到的脂肪酸符合生產(chǎn)生物柴油的標(biāo)準(zhǔn)。
3 結(jié)論
本文研究了不同濃度的CO2對(duì)WSE 中單針藻Monoraphidium sp. QLZ-3 生長(zhǎng)和油脂積累的影響。12%的CO2上調(diào)了rbcL 基因的表達(dá)量,加快了CO2的固定效率和多酚的吸收,同時(shí)上調(diào)了ACCase 和ME 的活性,下調(diào)了PEPC 的活性,提高了單針藻Monoraphidium sp.QLZ-3的生物量產(chǎn)率和油脂產(chǎn)率。綜上,CO2聯(lián)合WSE培養(yǎng)微藻,為微藻的工業(yè)化培養(yǎng)和核桃殼的綜合利用提供了新的思路。
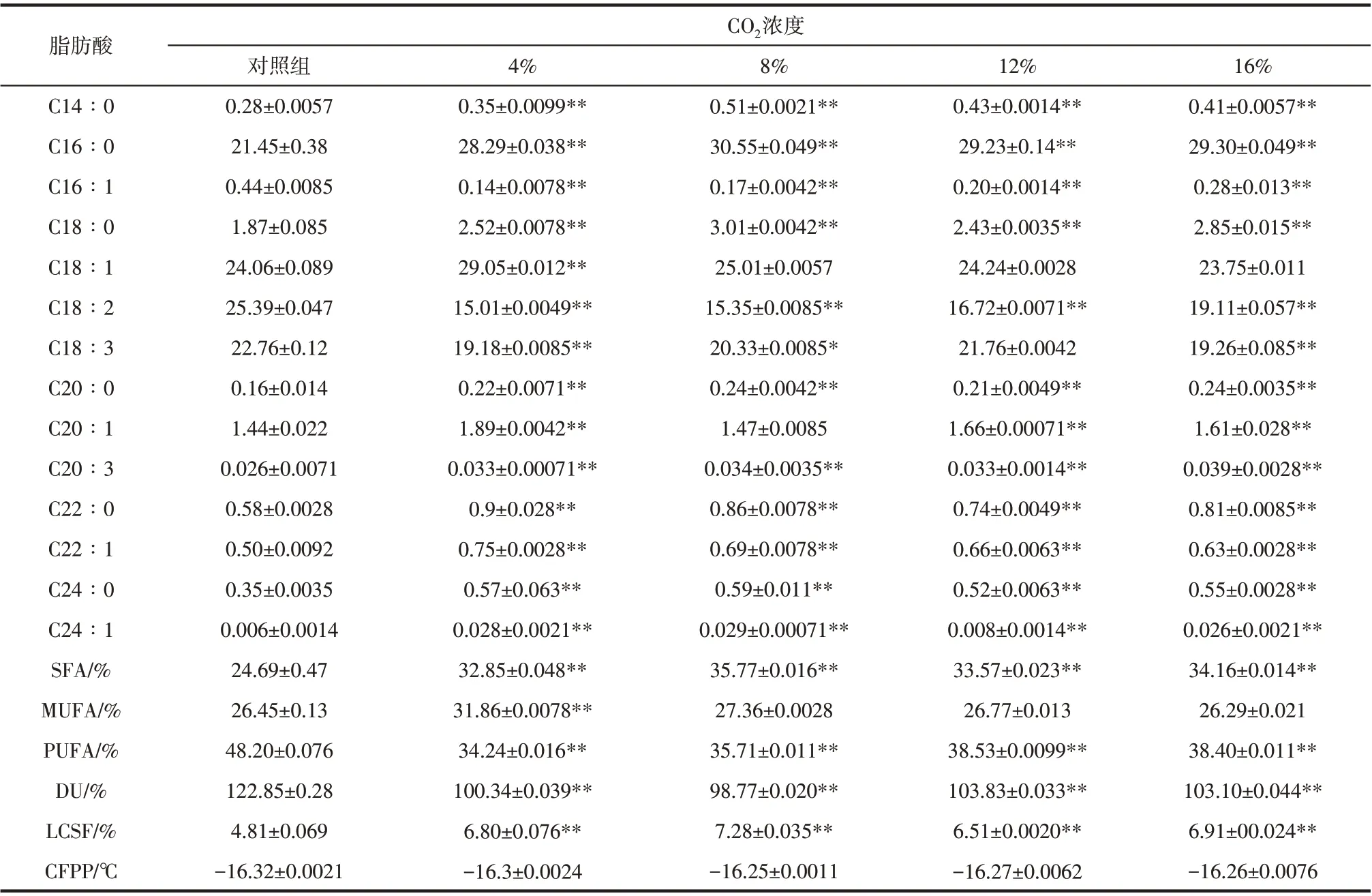
表2 不同CO2濃度對(duì)WSE中單針藻Monoraphidium sp.QLZ-3脂肪酸的組成的影響
猜你喜歡
食品工業(yè)(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫(yī)藥(2020年8期)2020-11-16 00:55:20
環(huán)境衛(wèi)生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語(yǔ)·新讀寫(xiě)(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國(guó)糧油學(xué)報(bào)(2019年4期)2019-07-12 09:06:38
食品與機(jī)械(2018年11期)2019-01-03 06:15:34
中國(guó)糧油學(xué)報(bào)(2018年12期)2018-01-26 13:32:04
