秈粳遺傳背景對水稻淀粉特性的影響
2020-04-21 09:00:26陶士博李鳴曉
北方水稻 2020年2期
關鍵詞:水稻
陶士博,李鳴曉,徐 銓
(1 遼寧省重要技術創新與研發基地建設工程中心,沈陽 110168;2 沈陽農業大學 水稻研究所,沈陽 110866)
水稻為全球超過50%的人口提供主食,是最重要的糧食作物之一(萬建民,2010)。隨著長期的馴化過程,水稻分化形成了不同的亞種,粳稻 (O.sativa ssp.japonica) 和秈稻 (O.sativa ssp.indica)[1-5]。 秈粳亞種在生物學特性上存在顯著差異,尤其是兩個亞種在水稻品質性狀上的差異尤為顯著。隨著人們生活水平的提高,水稻品質的提高也逐漸成了育種家和生物學家的目標, 秈粳之間的品質差異也成了研究的重中之重。 Nakamura Yasunori(2017)對水稻淀粉基因進行了全面的分析[6],我們選擇幾個重要的淀粉合成酶進行分析:水稻的蠟質基因(Wx)位于第6 染色體,其淀粉合成酶為存在秈粳特異性及突變型waxy (gbss1),且僅存在于水稻的支鏈淀粉中, 含有該基因的水稻外觀表現為純白色[7-9];SBE3 基因位于第 2 染色體且存在秈粳特異性, 其支鏈淀粉鏈長分布在6~13 范圍內, 有利于該淀粉合成酶的合成,在17~34 和大于39 范圍內,不利于該淀粉合成酶的合成, 含有該酶的水稻重量下降且稻米外觀品質存在堊白現象[10];FLO5 位于第 8 染色體,存在突變型,其基因型為 ss3a (e1)或者 floury-5,其支鏈淀粉鏈長分布在10~15 范圍內, 有利于該淀粉合成酶的合成,在6~9 和30~60 范圍內,不利于該淀粉合成酶的合成,外觀品質存在白核現象,且該酶的存在可以使 SSI 、GBSSI、AGPase 水平上升[11,12]; 另一重要的淀粉合成酶 UGP1 位于第 9 染色體,在外觀品質存在堊白現象,且可能雄性不育[13]。 為了進一步研究秈粳之間遺傳差異對水稻淀粉的影響,本文以重組自交系群體(RILs)為試驗材料,結合高通量測序,分析遺傳因素對水稻淀粉特性的影響, 同時也分析本文涉及的淀粉合成酶基因型, 并分別探究淀粉特性及秈型位點頻率(Fi)與品質性狀的相關性,加深對水稻淀粉的研究機理,從而選育出更適合人民需求的水稻品種,提供了遺傳學依據。
1 材料與方法
1.1 實驗材料
實驗選取了典型粳稻品種沈農265(SN265)與典型秈稻品種滬恢99(R99)雜交構建的重組自交系,重組自交系使用一粒傳法構建,自交10 代后得到155 個株系。 其中, 粳稻品種沈農 265(SN265)作為“中國超級稻育種”項目實施以來育成的第一個粳型超級稻品種,具有直立穗、穗短及抗逆性強的特點, 尤其是在稻瘟病的抗性表現上極為顯著(陳溫福,2004)。秈稻品種瀘恢 99(R99)作為秈稻的重要恢復系,具有彎曲穗、穗長的形態特點。 重組自交系和親本種植于四川省農業科學院水稻研究所德陽育種基地(N32°, E104°)。 155個株系按隨機區組試驗排列,每個株系種植3 行,每行種植 10 株, 栽植規格均為行距 30.0 cm,株距為 13.3 cm,并設 2 次重復。 栽培方法如下:在播種前分別測定了各地的土壤自然肥力, 按基礎肥力施用統一標準為每公頃150 kgN, 150 kgP,150 kgK 進行補施,并在移栽7 d 后施用返青肥,返青肥施用量為每公頃75 kgN。
1.2 水稻淀粉特性測定方法及數據分析
成熟期每個株系去除邊株后,取中間的20 株的稻穗,單獨進行脫粒。 稻谷脫粒后,在室溫環境下自然風干保存三個月后, 通過精米機將所收獲的稻谷碾磨成精米,將精米放入Foss 旋風磨中打粉,通過100 目篩子篩選,取精細米粉試樣留取備用。運用Nakamura 全波段紫外分光光度計法測定鏈長分布,使用日立(Hitachi)U3900 紫外分光光度計進行200~900 nm 波長下進行掃描。 每個株系的淀粉特性測定重復兩次。
1.3 秈型位點頻率(Fi)的劃分
秈型位點頻率,又稱Fi,我們定義親本粳稻品種沈農265(SN265)的秈型位點頻率為0,親本秈稻品種滬恢99(R99)的秈型位點頻率為1,試驗材料的秈型位點頻率指的是水稻株系偏向于秈稻親本即R99 的頻率, 篩選親本間有多態性、均勻分布12 條染色體的秈粳特異SNP 標記, 計算每個株系整個染色體組秈粳成分比例, 進而求得每個株系的秈型位點頻率, 從而更好地分析出其秈粳屬性與水稻淀粉特性間的關系, 對于秈型位點頻率 (Fi) 的具體劃分, 我們將秈型位點頻率(Fi) 小于 0.4 的劃分為偏粳型,0.4 到 0.6 為中間型,0.6 以上的為偏秈型。
1.4 QTL分析
QTL 區間作圖的方法最早提出于1989 年(Lander ES et al,1989),隨后王建康(2009)提出了大麥品種更為全面的數量性狀基因的完備區間的作圖方法, 對于水稻淀粉特性的QTL 分析,同樣適用,我們采用這種方法進行QTL 定位。 LOD閥值設為2.50 (不同環境下同時檢測到時LOD>2.40),當實際求得的 LOD 值大于 LOD 閥值時,就認為該區段存在1 個QTL,同時估算每個QTL 的加性效應值和貢獻率大小。
1.5 淀粉合成基因型分析
采用的是全基因重測序技術,對四個地區,每個地區155 個株系, 分析過程為先對已知的水稻基因組序列進行不同個體的基因組測序, 并在此基礎上, 對本次試驗材料個體或群體進行差異性分析。
1.6 數據處理
使用Microsoft Excel 2010 軟件處理數據和繪制圖表, 使用 SPSS 23.0 軟件統計分析。 四個地區的所有的樣品都測兩次重復, 數據結果使用2016 年的試驗數據用于下面的分析。
2 結果與分析
2.1 秈型位點頻率(Fi)與水稻淀粉特性的關系
我們將重組自交系群體通過秈粳特異 SNP標記得到的秈型位點頻率結果列于圖1。 由圖可知: 由于秈型位點頻率劃分界限導致部分株系早分界線處出現重疊,但我們可以看到,超過60%的株系處于中間型, 偏粳型的株系數略大于偏秈型的株系數。 由于秈型位點頻率(Fi)與淀粉特性之間相關性并不顯著, 于是我們進一步將不同地區按各自生態環境下, 用圖1 所列出的三種類型偏秈型、中間型、偏粳型作為SPSS 軟件多因素方差分析中的固定因子, 用淀粉特性的四個指標AAC、Fa、Fb3、RS 作為因變量(表1)。 結果表明秈型血緣并不顯著影響淀粉特性。

表1 秈粳類型間淀粉特性的差異
2.2 淀粉特性的QTL分析
我們依據水稻淀粉特性的四個指標AAC、Fa、Fb3及 RS 進行 QTL 定位, 得到以下數據 (表2),結果表明:對表觀直鏈淀粉(AAC)進行 QTL定位, 我們共找到2 個表觀直鏈淀粉 (AAC)的QTL, 位于第 1、7 染色體上,LOD 值介于 2.60~4.28, 加性效應值為-0.84~0.72, 單個貢獻率為11.58~15.55;對短支鏈淀粉(Fa)進行 QTL 定位,我們共找到1 個短支鏈淀粉(Fa)的QTL,位于第7 染 色體 上,LOD 值介于 2.92, 加性效應值為0.25,單個貢獻率為 7.70;對長分支支鏈淀粉(Fb3)進行QTL 定位,我們共找到5 個長分支支鏈淀粉(Fb3)的 QTL,位于第 1、3、4、5、6 染色體上,LOD值介于 3.43~41.11, 加性效應值為-1.25~1.45,單個貢獻率為 0.96~22.23; 對抗性淀粉 (RS) 進行QTL 定位, 我們共找到 2 個抗性淀粉 (RS)的QTL, 位于第 1、6、7 染色體上,LOD 值介于 2.80~3.23, 加性效應值為-0.19~0.15, 單個貢獻率為6.39%-12.68%。 第 1 染色體上的 2 位點能在表觀直鏈淀粉(AAC)、長分支支鏈淀粉(Fb3)、抗性淀粉(RS)都被檢測到,貢獻率在 1.69%~15.55%,加性效應為0.15~0.72,該位點可能是一個穩定表達的 QTL 區域(Block1835—Block1841),控制著表觀直鏈淀粉(AAC)、長分支支鏈淀粉(Fb3)、抗性淀粉(RS)含量;四川地區位于6 染色體上的62位點能在短支鏈淀粉(Fa)、長分支支鏈淀粉(Fb3)被檢測到, 貢獻率在 0.96%~7.70%, 加性效應為-0.30~0.25,該位點可能是一個穩定表達的QTL區域(Block13997—Block13994),控制著短支鏈淀粉(Fa)、長分支支鏈淀粉(Fb3)含量。
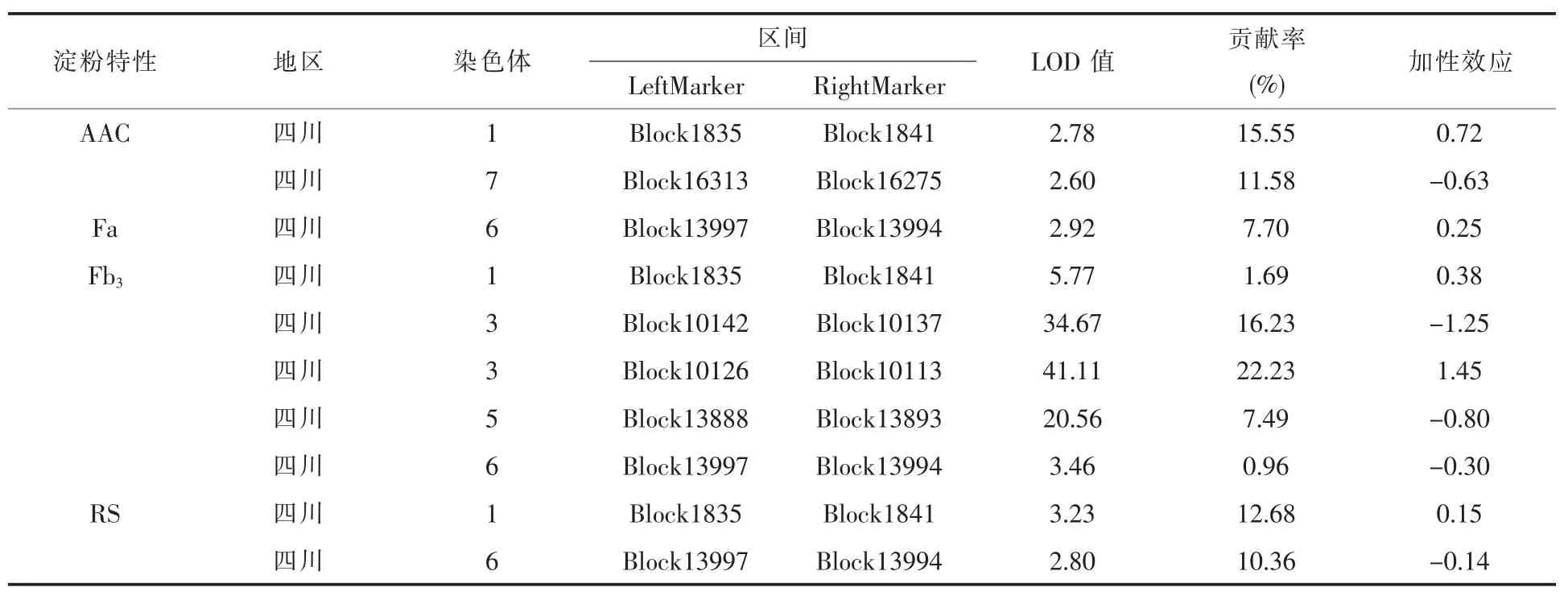
表2 淀粉特性的QTL 定位
2.3 淀粉合成相關酶的基因型分析
涉及水稻淀粉合成過程中的目標同工酶或調節因子共計28 個, 將這28 個淀粉合成過程中的目標同工酶或調節因子在本試驗中進行分析 (表3),結果表明在28 個涉及到水稻淀粉合成過程中的酶中,只有三個基因在雙親之間存在差異,它們分別是 SBE3、SSIIIa、UGP1,分別位于試驗材料的第2、8、9 染色體上。 針對于這三個淀粉合成相關酶的基因型, 我們將事先通過基因重測序得到的三個基因型的秈粳屬性以親本秈稻品種滬恢99(R99)和粳稻品種沈農 265(SN265)的淀粉特性為基準, 將除親本外的其他所有株系的淀粉特性同樣按照秈稻品種滬恢99(R99)淀粉特性接近、與粳稻品種沈農265(SN265)的淀粉特性接近以及與兩者都不接近的雜合型分為偏秈型Preindica、偏粳型 Pre-japonica、中間型 Intermediate,從而得到三種淀粉合成相關酶基因型秈粳屬性頻次分布圖(如圖2)。 結果表明:對于SBE3 基因型來說,偏粳型>偏秈型>中間型,SSIIIa 基因型偏秈型>偏粳型>中間型,UGP1 基因型偏粳型>偏秈型>中間型。為了更加深層次的研究在上述三種秈粳屬性下的三個淀粉基因型與水稻淀粉特性之間的關系, 我們將SPSS 22.0 處理下的多因素方差分析表匯總到表4。 結果表明:三個淀粉基因型與水稻淀粉特性之間均表現出極顯著的差異性,接下來單獨分析每個淀粉基因型, 我們發現SBE3基因型下, 不同秈粳屬性的株系與AAC、Fa、Fb3、RS 未達到顯著水平;SSIIIa 基因型下, 中間型的AAC 含量顯著高于偏秈型和偏粳型株系,中間型的Fa 含量顯著低于偏秈型和偏粳型株系,中間型的Fb3含量略高于偏秈型株系, 顯著高于偏粳型株系;UGP1 基因型下, 不同秈粳屬性的株系與AAC、Fa、Fb3、RS 未達到顯著水平。

表3 淀粉合成相關酶的基因型
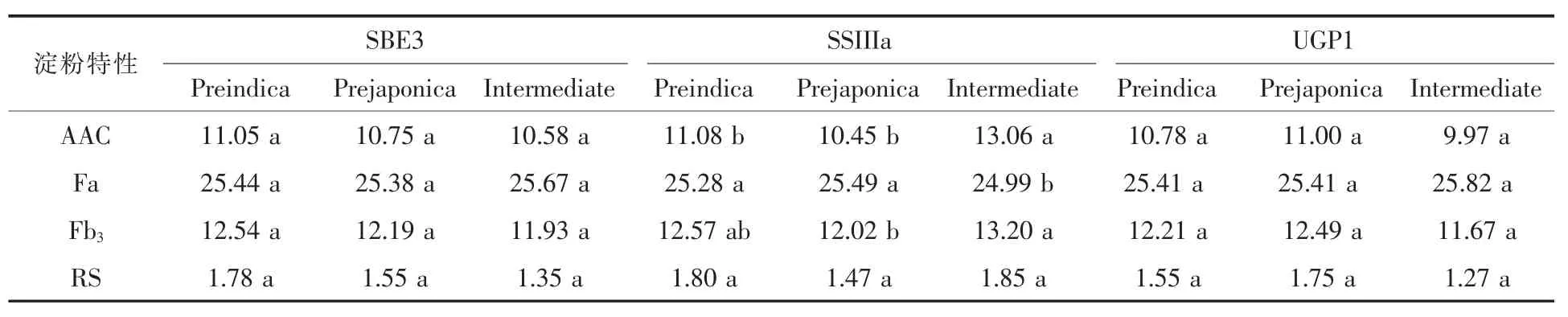
表4 不同淀粉基因型與淀粉特性差異性分析
3 討論
作為水稻精米胚乳中最主要的成分, 淀粉的研究日益深入。但目前來說,對于淀粉的研究多以不同生態環境背景或針對某一特定的淀粉基因型進行研究, 而涉及到水稻秈粳屬性與淀粉特性之間相關性的研究,目前還無相關研究。 我們發現,秈型頻率(Fi)與水稻淀粉特性之間的相關性雖有著一定的趨勢, 但其相關性并不顯著。 李修平等(2014)通過元分析的方法進一步分析了水稻直鏈淀粉含量的 QTL 及其連鎖圖譜標記[14], 孫春龍(2013)研究發現水稻抗性淀粉受基因加性效應控制,已知的蠟質基因(Wx)QTL 定位在水稻第6 染色體上[15]。為了更好的了解水稻淀粉特性,我們進行QTL 分析,發現位于第1 染色體上的2 位點能在表觀直鏈淀粉(AAC)、長分支支鏈淀粉(Fb3)、抗性淀粉 (RS) 都被檢測到, 貢獻率在 1.69%~15.55%,加性效應為 0.15~0.72,該位點可能是一個 穩 定 表 達 的 QTL 區 域 (Block1835—Block1841), 控制著四川地區的表觀直鏈淀粉(AAC)、長分支支鏈淀粉(Fb3)、抗性淀粉(RS)含量。 Chen M H et al(2008)認為在水稻淀粉特性中,表觀直鏈淀粉(AAC)是一個復雜的特征,受多種基因和環境的影響, 而最主要就是蠟質基因Wx[16]。 Wx 基因編碼顆粒結合的是淀粉合成酶 I(GBSSI),從而催化表觀直鏈淀粉(AAC)的合成。導致表觀直鏈淀粉(AAC)改變可能受到蠟質基因(Wx) 的表達的影響[17,18]。 除了淀粉特性中的AAC 外,其他淀粉特性指標 Fa、Fb3、RS 與淀粉合成酶之間的關系還有待進一步研究。 關于水稻淀粉基因型的分析,我們從28 個與淀粉合成過程中相關的淀粉合成酶進行篩選符合條件的三個重要淀粉合成酶基因SBE3、SSIIIa、UGP1 進行分析。其中SBE3、、UGP1 都表現出偏粳型株系略大于偏秈型,SSIIIa 表現出偏秈型略大于偏粳型,中間型表現不明顯。在此基礎上,我們更加需要了解在該分類下, 三種淀粉基因型與淀粉特性之間的差異性,結果顯示:三個淀粉基因型以親本淀粉特性為依據下, 與水稻淀粉特性之間均表現出極顯著的差異性,其中SBE3、UGP1 淀粉基因型,不同秈粳屬性的株系與 AAC、Fa、Fb3、RS 未達到顯著水平;SSIIIa 基因型, 中間型的AAC 含量顯著高于偏秈型和偏粳型株系,中間型的Fa 含量顯著低于偏秈型和偏粳型株系, 中間型的Fb3含量略高于偏秈型株系,顯著高于偏粳型株系,AAC、Fb3含量變化趨勢一致,與Fa 含量變化趨勢相反,再次印證了淀粉特性指標間的關系。 位于第8 號染色體上的SSIIIa 淀粉合成酶基因型對試驗材料中雜合型株系表觀直鏈淀粉 (AAC)、長分支支鏈淀粉(Fb3)含量有正向作用,對秈型株系和粳型株系短支鏈淀粉(Fa)含量有正向作用。 其作用機理還有待進一步研究。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
