木薯MebZIP7和MebZIP9基因的克隆及其轉錄活性分析
2020-03-17 06:06:00孟宇紅馬平安張圣奎王文泉
中國農(nóng)業(yè)大學學報 2020年2期
孟宇紅 陳 新 馬平安 宋 羽 張圣奎 王文泉*
(1.海南大學 熱帶農(nóng)林學院,海口 571101;2.中國熱帶農(nóng)業(yè)科學院 熱帶生物技術研究所,海口 571101;3.新疆農(nóng)業(yè)科學院農(nóng)作物品種資源研究所, 烏魯木齊 830091)
木薯(ManihotesculentaCrantz)起源于中南美洲亞馬遜熱帶雨林地區(qū),分屬大戟科(Euphorbiaceae)木薯屬(ManihotP. Mill.)多年生植物[1]。木薯、馬鈴薯和甘薯并稱世界三大薯類作物,所產(chǎn)淀粉廣泛用于食品和工業(yè)原料,是重要的糧食和經(jīng)濟作物[2]。木薯野生祖先種與栽培種的全基因組分析顯示:木薯栽培種的淀粉合成相關基因轉錄活性增強[3],而木薯塊根淀粉積累是一個復雜的生理代謝過程,會受到內源激素的調控,研究發(fā)現(xiàn)脫落酸(ABA)對木薯塊根淀粉積累起促進作用[4]。其他植物,如小麥內源激素ABA通過調節(jié)籽粒中蔗糖-淀粉代謝的關鍵酶活性,促進籽粒灌漿[5],外源ABA處理水稻種子后籽粒千粒重增加,產(chǎn)量增加[6]。
堿性亮氨酸拉鏈(bZIP)是家族成員眾多的一類轉錄因子。bZIP轉錄因子有高度保守的bZIP區(qū),由一個堿性區(qū)和一個亮氨酸拉鏈組成[7]。依據(jù)bZIP相似的堿性區(qū)和附加的保守序列將模式植物擬南芥的bZIP基因劃分為10個亞族,分別為A-I和S亞族,其中 A亞族與ABA相關,屬于脫落酸響應元件結合因子(ABF)[8]。ABF可以反式激活在啟動子區(qū)含有脫落酸響應元件(ABA response element,ABRE)的效應基因,它的表達受ABA和各種脅迫誘導[9]。ABF參與ABA信號通路,ABA結合PYL后破壞了SnRK2蛋白激酶和PP2C的相互作用,進而激活SnRK2蛋白激酶使ABF磷酸化,從而調節(jié)響應ABA信號相關基因的表達[10-12]。擬南芥ABF2過表達植株中SuSy1(Sucrose synthase1)的表達水平低于野生型,高濃度鹽處理后SuSy1在轉基因植株中的轉錄水平高[13]。
基因組研究表明木薯堿性亮氨酸拉鏈(MebZIP)同樣是一個大的轉錄因子家族[14],目前對其研究報道相對較少。本研究針對bZIP A亞族基因:MebZIP7和MebZIP9,通過測定它們在淀粉積累效率不同木薯野生和栽培品種中的轉錄活性,以及在外源ABA誘導下的表達模式,分析MebZIP成員是否參與木薯塊根淀粉合成代謝調節(jié),旨在為深入研究bZIP在ABA參與木薯塊根淀粉調控中的作用提供一些試驗依據(jù)。
1 材料與方法
1.1 試驗材料
試驗選用栽培木薯KU50、SC8、Arg7和野生亞種W14(Manihotesculenta.ssp.flabellifolia),3個 栽培種的塊根淀粉含量高于野生型。KU50和W14用于木薯全基因組測序。SC8是中國熱帶農(nóng)業(yè)科學院培育的代表性品種,用于ABA處理的組培苗。種植在海南澄邁試驗基地,在種植后90(塊根形成期)、180(塊根膨大期)和210 d(塊根成熟期)分別取塊根及功能葉用于分析。
選擇苗齡45 d,長勢均一的SC8組培苗進行ABA處理。每個培養(yǎng)瓶中加入1.0 mL 100 μmol/L的ABA溶液,使其均勻覆蓋于培養(yǎng)基表面。分別處理1、6、12和24 h,取處理后的葉片和塊根做后續(xù)試驗。每個處理3次重復和1個對照(未經(jīng)ABA處理)。3個重復的混合樣放入液氮中速凍,-80 ℃保存待用。
1.2 10個MebZIP基因的確定及實時熒光定量表達分析
在轉錄因子數(shù)據(jù)庫(http://planttfdb.cbi.pku.edu.cn/)中搜索到104個木薯的bZIP轉錄因子,結合本實驗室已有的木薯轉錄組數(shù)據(jù)[3],挑選栽培型與野生型塊根中相對表達量比值≥2的MebZIP基因,初步篩選到10個MebZIP基因(表1)。將初步篩選到的10個MebZIP基因,根據(jù)基因ID號依次命名為MebZIP1~MebZIP10。
取木薯栽培種KU50、Arg7、SC8和野生型W14的塊根及功能葉,利用實時定量PCR(qPCR)檢測10個MebZIP在不同品種和不同組織部位的差異表達。用Primer- BLAST(https://www.ncbi.nlm.nih.gov/tools/primer-blast/)設計定量引物并檢測引物的特異性,qPCR引物序列見表2。使用木薯Manes.02G137500作為內參基因[14]。用Mx3000P QPCR儀,大連寶生物公司的PowerUpTMSYBRTMGreen Master Mix試劑進行試驗。qPCR程序: 95 ℃預變性2 min,95 ℃變性15 s,60 ℃退火1 min,72 ℃延伸20 s,40個循環(huán),溶解曲線95 ℃ 1 min,55 ℃ 30 s,95 ℃ 30 s,并用2-ΔΔct法整理分析數(shù)據(jù)[15]。

表1 10個 MebZIP在栽培種KU50和野生型W14塊根中的轉錄本數(shù)量及比值Table 1 The transcription and ratio of ten MebZIP in the cultivated species KU50 and wild-type W14

表2 10個MebZIP基因實時定量分析所用引物序列Table 2 Primer sequences of ten MebZIP genes for qPCR
1.3 植物總RNA提取及單鏈cDNA的合成
使用RNA plant Plus Reagent植物總RNA提取試劑,購于天根(北京)公司。進行木薯總RNA的提取。RNA質量及濃度通過瓊脂糖凝膠電泳和微量紫外分光光度計檢測。使用Prime ScriptTMRT reagent Kit with gDNA Eraser反轉錄試劑盒購于寶生物(大連)公司合成單鏈cDNA,并用木薯通用Actin引物檢測第一鏈cDNA。RNA保存于-80 ℃,cDNA保存于-20 ℃。
1.4 10個木薯MebZIP與127個擬南芥AtbZIP氨基酸序列比對
在轉錄因子數(shù)據(jù)庫(http://planttfdb.cbi.pku.edu.cn/)中搜索擬南芥bZIP基因,得到127個bZIP。下載擬南芥所有bZIP基因的氨基酸序列與木薯候選的10個bZIP基因的氨基酸序列經(jīng)CLUSTAL W進行序列比對,之后使用MEGA 5.0構建系統(tǒng)進化樹。進化樹的構建使用NJ(Neighbor-Joining)法,并經(jīng)1 000次Bootstrap統(tǒng)計檢測。
1.5 MebZIP7和MebZIP9基因的克隆及MebZIP-GFP載體的構建
從木薯基因組數(shù)據(jù)庫(https://phytozome.jgi.doe.gov)中搜索目的基因序列(從10個MebZIP中選擇2個MebZIP),并用Primer-BLAST設計特異性擴增引物,引物序列信息:MebZIP7上游引物(5′TGTTCATGGTTGTTAGGAATTGGT 3′)和下游引物(5′TCCTTGCACCACACAAATGG 3′);MebZIP9上游引物(5′AAGAAACTGAACA-GGAACGTCTG 3′)和下游引物(5′CTTCCCTT-CCACTGCTATACCT 3′)。用大連寶生物公司的Primer STARMax DNA Polymerase,以KU50塊根cDNA為模板進行基因擴增。PCR程序:98 ℃預變性5 s,98 ℃變性10 s,55 ℃退火5 s,72 ℃延伸15 s,30個循環(huán)后72 ℃延伸5 s,用MEGA5.0進行測序后序列比對。
MebZIP基因插入pCAMBIA1302載體中,所用引物見表3。通過激光掃描共聚焦顯微鏡觀察MebZIP-GFP的融合蛋白亞細胞定位。

表3 構建MebZIP-GFP載體所用引物Table 3 Primer sequences for constructing MebZIP-GFP plasmid
2 結果與分析
2.1 木薯MebZIPs在木薯栽培種和野生型中具有不同的表達譜
熒光定量表達分析結果,MebZIP1~MebZIP10在栽培品種KU50塊根中的相對表達量均高于野生亞種W14,表達量比值在1.4~185.0倍,其中MebZIP9最高,MebZIP1最低,MebZIP7達到2.3倍,6個成員如MebZIP2、MebZIP3、MebZIP4、MebZIP7、MebZIP8和MebZIP9表達差異達到顯著和極顯著水平(圖1(a))。 而在功能葉中,MebZIP4、MebZIP9和MebZIP10在栽培品種的相對表達量高于野生型,相差倍數(shù)>2(圖1(b))。特別是MebZIP9在塊根和功能葉中的相對表達量栽培品種均顯著高于野生型,倍數(shù)在39~185。表明塊根淀粉積累效率高的栽培品種塊根中存在MebZIP成員的優(yōu)勢表達,且在功能葉中也有少數(shù)成員高表達。
2.2 系統(tǒng)分類表明MebZIP7和MebZIP9與ABA信號通路相關
從轉錄因子數(shù)據(jù)庫(http://planttfdb.cbi.pku.edu.cn/)中搜索到擬南芥bZIP 127個,10個MebZIP與AtbZIP的蛋白質序列構建進化樹。bZIP轉錄因子家族總共劃分為10個亞族,分別為A、B、C、D、E、F、G、H、I和S亞族,而木薯MebZIP5、MebZIP7和MebZIP9屬于bZIP家族的A亞族,MebZIP2、MebZIP3和MebZIP10屬于S亞族,MebZIP1和MebZIP4屬于G亞族,MebZIP8屬于C亞族,MebZIP6屬于B亞族(圖2)。A亞族在 ABA信號轉導中起重要作用,C亞族可能調控貯藏蛋白在擬南芥胚乳里的表達,G亞族在光信號轉導和種子成熟過程中起作用,S亞族在蔗糖信號以及響應脅迫中起作用等。系統(tǒng)分類結果證明,MebZIP7和MebZIP9屬于A亞族,歸于ABF類,是ABA信號通路成員。
2.3 MebZIP7和MebZIP9定位于細胞核內
激光共聚焦顯微鏡下觀察DAPI染色后的洋蔥表皮細胞,MebZIP7-GFP和MebZIP9-GFP表達的蛋白均定位于細胞核,在細胞壁及細胞質內無綠色熒光,而對照試驗CaMV35s::GFP在整個洋蔥表皮細胞都有GFP分布(圖3)。據(jù)此,可以確定MebZIP7和MebZIP9均具有細胞核定位特征。bZIP是一個大的轉錄因子家族,該試驗證明木薯中這2個成員屬于轉錄因子功能蛋白。

KU50,SC8和Arg7是栽培木薯;W14是野生木薯。KU50,SC8 and Arg7 are cultivated cassava. W14 is wild cassava.同組柱形圖不同小寫字母表示品種間表達差異顯著(P<0.05)Different letters above the column indicate significant differences in expression between the two varieties of cassava.圖1 10個MebZIP基因在栽培和野生型塊根(a)與葉片(b)中的表達差異Fig.1 Tissue-specific expression of ten MebZIP genes in root (a) and leaves (b) of cultivated and wild species

圖2 構建10個MebZIP系統(tǒng)發(fā)育樹Fig.2 Constructing phylogenetic tree of ten MebZIPs

綠色,GFP是綠色熒光蛋白,激光共聚焦顯微鏡下發(fā)出綠色熒光;藍色,DAPI染液染色,激光共聚焦顯微鏡下發(fā)出藍色熒光。標尺:20 μmGreen,GFP are green fluorescent proteins,which emit green fluorescence under laser confocal microscopy; Blue,DAPI dye staining and emits blue fluorescence under laser confocal microscopy. Bar: 20 μm圖3 MebZIP7和MebZIP9在洋蔥表面亞細胞定位Fig.3 MebZIP7 and MebZIP9 locate in the nucleus of onion surface cell
2.4 MebZIP7和MebZIP9受外源ABA誘導上調表達
ABA處理木薯后1 h,MebZIP7被強烈誘導表達且相對表達量達到峰值,此時根中和葉片的相對表達量分別是對照的10.7和4.5倍;MebZIP7的相對表達量隨處理時間的增加而逐漸降低,24 h時與初始水平一致(圖4)。MebZIP9同樣被ABA誘導上調,1 h根中的相對表達量達到峰值,是對照的6.3倍;在葉片中的相對表達量在6 h達到最高,是對照的4.5倍;MebZIP9在ABA處理下的相對表達量先升高后下降,24 h根中的相對表達量低于初始水平(圖5)。盡管在幼苗培養(yǎng)條件下與自然生長發(fā)育條件有很大差異,仍然可以捕捉到木薯根和葉片中MebZIP7和MebZIP9受到外源ABA信號誘導而增強表達,說明2個bZIP成員確實參與ABA信號途徑。

1L和1R:葉片,塊根ABA處理1 h;6L和6R: 葉片,塊根ABA處理6 h;12L和12R:葉片,塊根ABA處理12 h;24和24R:葉片,塊根ABA處理24 h。下同。1L and 1R, leaf and root are treated by ABA for 1 h; 6L and 6R, leaf and root are treated by ABA for 6 h; 12L and 12R,leaf and root are treated by ABA for 12 h; 24L and 24R, leaf and root are treated by ABA for 24 h. The same below. 圖4 MebZIP7受ABA誘導在葉片(a)和塊根(b)中的表達分析Fig.4 The expression profiles of MebZIP7 induced by ABA in leaf (a) and storage root (b)

圖5 MebZIP9受ABA誘導在葉片(a)和塊根(b)中的表達分析Fig.5 Expression profiles of MebZIP9 induced by ABA in leaf (a) and storage root (b)
2.5 MebZIP9在栽培木薯中的轉錄活性高于野生型
對于另一個基因MebZIP7在栽培品種和野生型不同發(fā)育時期塊根及葉片中均有表達,表達差異不能夠明確區(qū)別。如在塊根發(fā)育90 d,180 d時KU50、SC8和Arg7中表達量高于W14,而在210 d時差異不明顯;葉片中,90 d時W14的表達量高于幾個栽培品種,其他時間點表達量差異不顯著(圖6)。MebZIP9在栽培與野生型的塊根和葉片中表現(xiàn)出完全不同的表達模式。在野生型W14整個生長期的塊根和葉片中相對表達量均很低,而在不同木薯栽培品種不同發(fā)育時期的塊根和葉片中均保持較高表達量,塊根中表達量更高一些。其表達量總體有前期高后期下降的趨勢,如在KU50塊根形成期相對表達量最高,在塊根膨大期和塊根成熟期的相對表達量相同,最高值與最低值相差2倍;在SC8塊根形成期和塊根膨大期的表達量相近,是塊根成熟期的1.4倍;在Arg7 塊根膨大期相對表達量高,是塊根形成期和成熟期的3.6倍(圖7)。此外,MebZIP7比MebZIP9在塊根和葉片發(fā)育不同時期整體表達量更高。
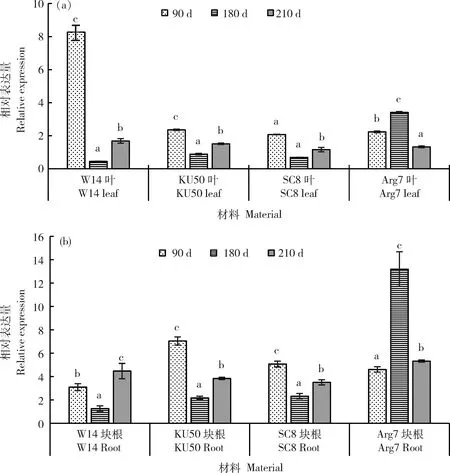
同組不同小寫字母表示表達量差異顯著(P<0.05)系指每組材料間表達差異顯著性。下同。The marked in the figure refers to the significance of difference in expression quantity between each group of materials. The same below.圖6 MebZIP7在木薯栽培型及野生型葉片(a)和塊根(b)中的差異表達Fig.6 Spatio-temporal expression profiles of MebZIP7 in leaf (a) and root (b) of four cassava varieties

圖7 MebZIP9在木薯栽培品型及野生型葉片(a)和塊根(b)中的差異表達Fig.7 Spatio-temporal expression profiles of MebZIP9 in leaf (a) and root (b) of four cassava varieties
3 討 論
3.1 木薯bZIP成員在塊根中有優(yōu)勢表達,可能與其淀粉積累效率有關
MebZIP9在栽培木薯塊根中的轉錄活性顯著高于野生型,MebZIP9在同一木薯同時期塊根和葉片中的表達水平低于MebZIP7。
MebZIP7在栽培種塊根形成期(90 d)和塊根膨大期(180 d)塊根中的相對表達量都顯著地高于野生型,MebZIP9在栽培種整個生長期塊根中的相對表達量都顯著的高于野生型,特別是MebZIP9在W14塊根中幾乎不表達。栽培木薯淀粉含量顯著高于野生型[3],木薯栽培品種淀粉含量(干重)為28%~32%,野生型W14淀粉含量是3%~5%。綜上所述,推測MebZIP7和MebZIP9可能參與木薯塊根淀粉合成。現(xiàn)有研究發(fā)現(xiàn):bZIP可以調節(jié)淀粉合成,例如,水稻C亞族的OsbZIP58在胚乳中高表達,其蛋白可直接與6個淀粉合成相關基因(OsAGPL3,Wx,OsSSIIa,SBE1,OsBEIIb和ISA2)的啟動子區(qū)結合并調控它們的表達活性,是水稻籽粒中淀粉合成的重要調節(jié)因子[16]。玉米I亞族的ZmbZIP91可與玉米淀粉合成相關基因啟動子的ACTCAT元件結合,進而調節(jié)玉米淀粉相關基因的表達[17]。據(jù)此推測MebZIP可能調節(jié)木薯塊根淀粉合成相關基因的表達或淀粉合成酶的活性。
3.2 木薯bZIP成員受外源ABA誘導表達
高濃度的ABA抑制水稻籽粒灌漿期淀粉合成基因的表達和淀粉合成酶的酶活,導致籽粒充實率降低[18],而適中濃度ABA能增強水稻籽粒灌漿期SuSy的活性[19]。擬南芥葉片中β-淀粉酶1(BAM1)和α-淀粉酶3(AMY3)受ABA誘導表達,導致BAM1酶活提高,葉片淀粉含量降低[20]。MebZIP7和MebZIP9受ABA誘導表達,原因可能是:一方面,MebZIP7、MebZIP9是ABF(ABRE),ABF是ABA信號通路中的成員[21- 22],ABA結合PYL后破壞了SnRK2蛋白激酶和PP2C的相互作用,進而激活SnRK2蛋白激酶使ABF磷酸化,即ABF (MebZIP7和MebZIP9)表達量增加,從而調節(jié)響應ABA信號相關基因的表達。另一方面相關文獻報道[13-14]bZIP響應各種非生物脅迫,植物做出一系列抵抗外源ABA脅迫的機制,響應ABA的相關基因轉錄活性增強,因而與其上游啟動子結合的MebZIP轉錄因子的表達量增高。綜上推測栽培木薯中MebZIP受適量ABA誘導高表達并與淀粉合成相關基因的啟動子區(qū)結合,調節(jié)塊根淀粉合成。
此外,轉錄因子在植物里的表達水平很低[23],本研究選用管家基因Manes.02G061900[14],其在木薯各組織部位及脅迫處理后的表達水平都很穩(wěn)定,而且比常用的木薯Actin基因Manes.12G150500低1.3倍,使用Manes.02G061900做內參基因,便于對表達量低的轉錄因子進行分析。MebZIP7和MebZIP9在栽培木薯塊根中的表達水平顯著高于野生型且受ABA誘導表達,為后續(xù)分析MebZIP在溝通ABA信號通路和塊根淀粉合成途徑中的作用提供有效借鑒。
