以蛋白翻譯后修飾的視角探究脂肪肝的發病機制
2020-03-11 03:25:58張嘉洸盛亮
藥學研究 2020年1期
張嘉洸,盛亮
(南京醫科大學基礎醫學院藥理系,江蘇 南京 211166)
脂肪肝問題已經成為遍及全球的健康問題,由脂肪肝引起的一系列機體代謝紊亂嚴重危害了人體的健康和生活質量。脂肪肝是非酒精性脂肪性肝炎、肝硬化、肝細胞癌和肝功能衰竭的既定危險因素。它還與血脂異常、2型糖尿病和高血壓的發展風險增加有關[1]。而脂肪肝的發生,主要是由于脂質的沉積過多引起的肝臟脂肪變性,要研究脂肪肝的發病機制和治療措施,就要從與脂質代謝相關的方面入手。當肝臟組織中脂肪酸(FA)輸入的速率[攝取和合成以及隨后酯化成甘油三酯(TG)]大于FA輸出(氧化和分泌)的速率時,就會發生脂肪變性[2]。而導致肝臟脂質增多的因素包括了4個方面:①過多游離脂肪酸的攝取;②增加從頭脂肪合成;③降低脂肪的氧化;④減少脂肪酸的出口[3]。每個方面的代謝過程都會受到多種酶和蛋白質的調控,對這類酶和蛋白質的翻譯后修飾,可能會對脂肪肝的發生發展有著一定的影響。本文主要從4個方面總結了多個與脂質代謝相關的蛋白翻譯后修飾的方式和對脂質代謝過程的影響。
1 脂肪的分解
在正常人體組織中,脂類絕大部分以TG的形式儲存在脂肪組織中,無論是要進行脂肪的氧化分解或是內源性脂肪酸的合成,都需要進行脂肪動員,將TG分解為游離脂肪酸[3]。TG的水解是一系列有序調控的過程,TG首先被脂肪甘油三酯脂肪酶(ATGL),激素敏感性脂肪酶(HSL)水解成甘油二酯(DG)和游離脂肪酸(FFA)[4],其次,甘油二酯被HSL水解成甘油一酯(MG)和FFA,最后甘油一酯被單線甘油酯酶(MGL)水解成甘油和FFA(見圖1)。
若脂質的分解過程紊亂,過多的脂質分解會導致內源性脂肪酸的合成和脂質的氧化增多,導致機體內的脂質堆積和氧化損傷。

圖1 脂肪的水解
1.1 脂肪甘油三酯脂肪酶(ATGL) ATGL是在細胞內脂肪分解過程中使TG儲存釋放脂肪酸的關鍵酶,提供肝臟組織所需要攝取的游離脂肪酸,主要參與TG分解為DG的過程,其能力強于HSL對脂質的分解作用。同時ATGL也是脂解的限速酶,ATGL的缺乏可能會造成細胞內脂質的空泡化沉積,尤其是在近曲小管處。嚴重的脂質沉積會帶來細胞凋亡等問題,導致腎臟纖維化和功能障礙[5]。
ATGL的修飾方式主要為:①E3泛素連接酶COP1(也稱為RFWD2)與ATGL的共識VP基序結合并通過主要在賴氨酸100殘基上K-48連接的多泛素化作用將其靶向蛋白酶體降解,降低ATGL的活性和效應,引起脂質的堆積[6];②TSH(促甲狀腺激素)與TSHR(促甲狀腺激素受體)結合后,激活蛋白激酶A(PKA),PKA結合ATGL的絲氨酸406殘基,抑制了ATGL的活性[7];③酰基輔酶A(LCA)直接與ATGL N-末端patatin樣磷脂酶結構域(PNPLA)結合,抑制了ATGL活性[8]。
1.2 激素敏感脂肪酶(HSL) HSL基因位于染色體19Q 13.3,HSL由脂肪組織分泌,也在胰腺、睪丸、肌肉、腎上腺等組織發現表達。HSL是一種細胞內的中性脂肪酶,特異性比較弱,能水解甘油三酯、甘油二酯、甘油一酯和膽甾烯基酯,卻沒有水解磷脂酶活性。伴隨HSL的基因敲除,脂肪水解能力被破壞,脂質合成和脂肪代謝能力明顯下降,這說明了HSL在脂肪的合成及分解代謝中起著無可替代的作用。與ATGL相比,ATGL催化TG底物的能力強于催化DG底物的能力,而HSL催化DG底物的能力10倍于催化TG底物的能力,ATGL和HSL共同催化哺乳動物儲存在脂肪組織內的TG[9]。HSL的缺乏可導致年齡依賴性的肝臟脂肪變性,誘導脂肪肝的產生[10]。
HSL具有兩個磷酸化位點。站點1被稱為監管站點,位于HSL蛋白序列中Ser-563的位置,被認為在HSL的激活中起關鍵作用。該位點被環AMP依賴性蛋白激酶和糖原合成酶激酶-4磷酸化。同時還受到激素的調控,腎上腺素通過cAMP級聯反應PKA磷酸化HSL,而胰島素水解cAMP降低PKA的含量來降低HSL的磷酸化,降低其對于脂質的水解作用。站點2稱為基礎站點,該位點置于HSL蛋白中Ser-565的位置,被兩種蛋白激酶磷酸化,即Ca2+/鈣調蛋白依賴性激酶Ⅱ和AMP激活蛋白激酶。同時,Ser-659和Ser-660也是負責激活HSL的主要PKA磷酸化位點。此外,PKC可通過細胞外信號調節激酶(ERK)依賴性的途徑磷酸化HSL的Ser-600并增加脂肪細胞中的HSL活性。
關于HSL的去磷酸化,Ser-563被B56alpha/PP2A(蛋白磷酸酶2A)和PP2C有效去磷酸化,而Ser-565主要被PP2A去磷酸化[11]。
2 游離脂肪酸的攝取
人體內的游離脂肪酸可來自內源性的脂肪合成,甘油三酯的水解和外源性的脂肪攝入。游離脂肪酸經肝實質細胞攝取后,可進行合成脂肪和脂肪氧化兩種步驟,為機體儲存能量或是提供機體所需要的能量消耗。無論是進行那個步驟,游離脂肪酸都要先進行活化,變成脂肪酰基輔酶A才可參與下一步的反應,其中需要長鏈酰基輔酶A合成酶(ACSL1)等酶的作用(見圖2)。
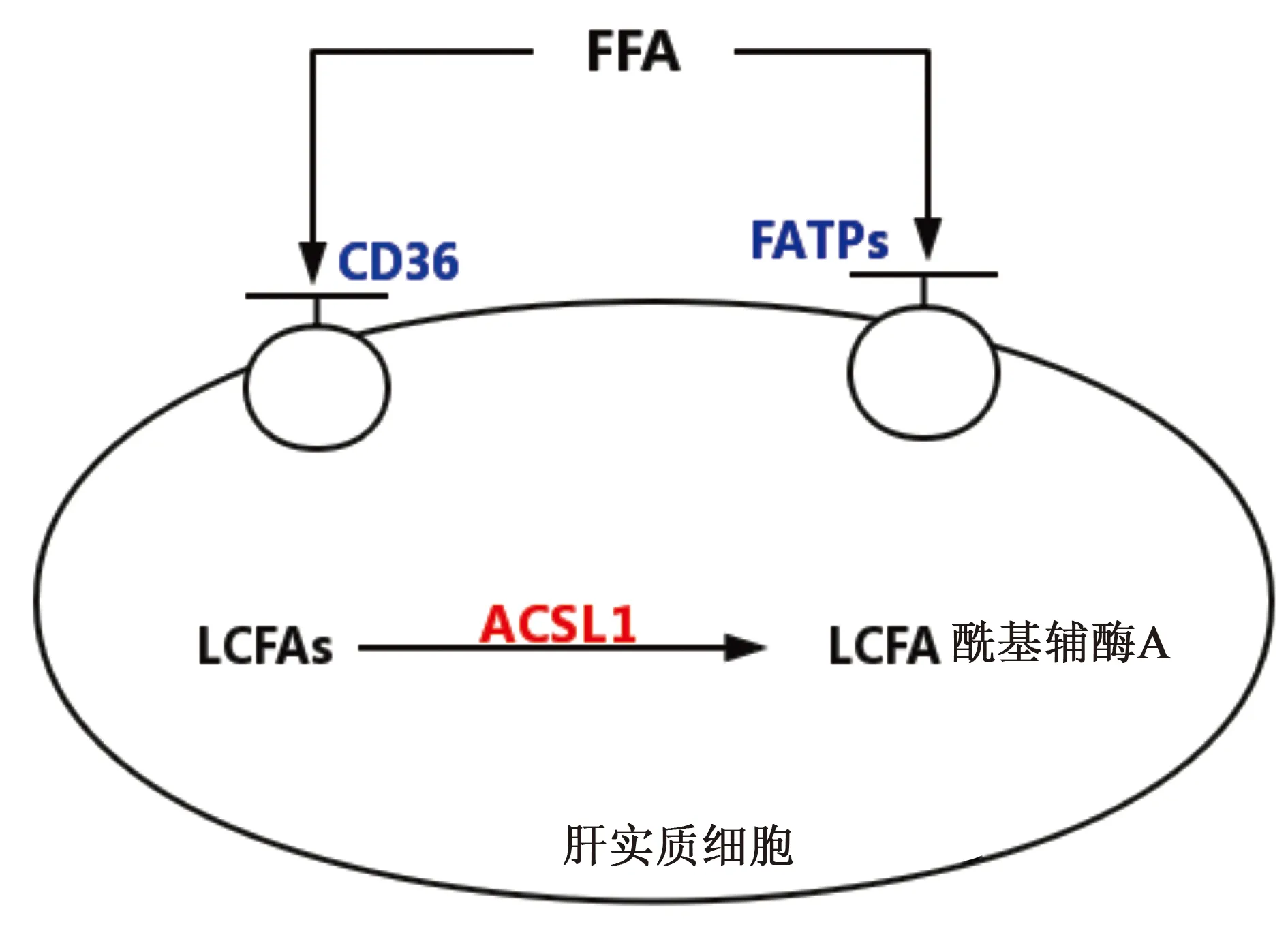
圖2 游離脂肪酸的攝取
2.1 CD36 CD36是清道夫受體,屬于多功能蛋白,是多種配體的共同受體。其在高親和力組織攝取長鏈脂肪酸(FAs)中起重要作用,將游離脂肪酸轉運到細胞內,供細胞利用[12]。CD36的活性若受到抑制會減少極低密度脂蛋白(VLDL)的輸出,可能是減輕脂肪肝的一種可行的方法[13]。
CD36受到的翻譯后修飾的主要方式為:①PKC與PKA可分別磷酸化CD36的Thr-92和Ser-237,抑制了CD36對脂質的攝取。而腸道的堿性磷酸酶gIAP可使CD36去磷酸化,使得其攝取脂質的能力提高[14];②CD36還受泛素化的調節,CD36是Parkin(E3連接酶)的靶點,Parkin作用于CD36 C末端的賴氨酸,使CD36水平升高,穩定性增加[15];③FA攝取和信號傳導中的CD36可被長鏈FA的磺基-N油酸羥基琥珀酸亞胺(NHS)酯不可逆地抑制。磺酸-N-琥珀酰亞胺油酸酯(sso)為其中一種,sso可作用于CD36的Lys-164,使CD36乙酰化提高它攝取脂質的能力[16]。
2.2 脂肪酸轉運蛋白1(FATP1) FATP1是FATP家族成員,主要分布在脂肪組織和肌肉組織中,同時也分布在肝臟細胞內。它可增強細胞對長鏈脂肪酸(LCFA)的攝取[17]。它對脂肪酸的轉運能力較CD36弱,但是仍是人體內主要的脂肪酸轉運蛋白,若活性降低即可減少組織對脂肪酸的攝取,減少細胞內脂肪的沉積。同時,FATP1還與視網膜有關,FATP1的缺乏會影響視網膜的光反應和暗適應[18]。
雷帕霉素復合物1(mTORC1)和p70核糖體蛋白S6激酶1(S6K1)軸可激活導致肥胖和衰老的代謝途徑,其中就包括對FATP1的激活。mTORC1-S6K1在Ser999處對谷氨酰 - 脯氨酰tRNA合成酶(EPRS)的磷酸化,誘導其從氨酰基tRNA多轉錄酶復合物中釋放,磷酸化EPRS結合FATP1,激活FATP1,誘導其轉運至質膜和長鏈脂肪酸攝取[19]。
2.3 長鏈酰基輔酶A合成酶1(ACSL1) ACSL1在脂肪酸代謝中起關鍵作用,是長鏈脂肪酸硫酯化成酰基輔酶A衍生物所必需的。長鏈脂肪酸(LCFAs)轉運到細胞內后,無論是進行合成代謝還是分解代謝,都需要進行活化,即在ACSL1的作用下,轉化為長鏈脂肪酰基輔酶A,所以ACSL1是長鏈脂肪酸進入代謝的第一步反應的催化酶。如果ACSL1的活性或水平急劇變化,長鏈脂肪酰基輔酶A增多,在肝臟中蓄積的TG和氧化產生的自由基也增多,將導致脂肪肝的加重和肝臟的氧化損傷。但肝臟中ACSL1的特異性缺失并不會影響肝臟中的脂質水平[20]。
所有入選患者由專業的視光醫師給予驗配試戴,最后確定鏡片參數。戴鏡時間保證每晚8~10 h,戴鏡后1個月、3個月和6個月復查。本項研究遵循赫爾辛基宣言和臨床研究的倫理標準和規范,向入選患者交代檢查流程,參與研究的青少年患者及其監護人均知情同意。所有檢查由同一名經驗豐富的醫師操作。
ACSL1的翻譯后修飾主要為磷酸化和乙酰化,AMPK可作用于ACSL1的Ser-278,磷酸化ACSL1抑制其活性,而SIRT3可使ACSL1的Lys-285脫乙酰化而增強其活性[21]。
3 脂肪的從頭合成
脂肪酸在體內的合成過程復雜,受到多種因子的調控,某個酶或細胞因子的活性及水平改變都會影響到內源性的脂質合成。
脂質的從頭合成需要的原料為乙酰輔酶A,來源于葡萄糖的代謝。葡萄糖通過糖酵解水解成丙酮酸,丙酮酸進入線粒體產生乙酰輔酶A。乙酰輔酶A通過檸檬酸合成酶與草酰乙酸結合形成檸檬酸鹽。檸檬酸鹽被輸出到細胞質中并通過ATP-檸檬酸裂解酶(ACLY)分裂成乙酰輔酶A和草酰乙酸。在細胞質中,乙酰輔酶A被乙酰輔酶A羧化酶(ACC)羧化形成丙二酰輔酶A。丙二酰輔酶A和NADPH均用作于乙酰輔酶A,通過脂肪酸合成酶(FASN)合成棕櫚酸(16碳脂肪酸)。棕櫚酸被內質網(ER)中的脂酰輔酶A延伸酶(Elov1)家族成員延長以產生長鏈脂肪酸(LCFA)(> 16碳鏈)。LCFA通過硬脂酰輔酶A去飽和酶(SCDs),形成單不飽和和多不飽和的LCFA。不飽和脂肪酸再在mtGPAT1和Dgat1/2的催化下與三磷酸甘油酯化形成三酰基甘油(TAG)[20](見圖3)。
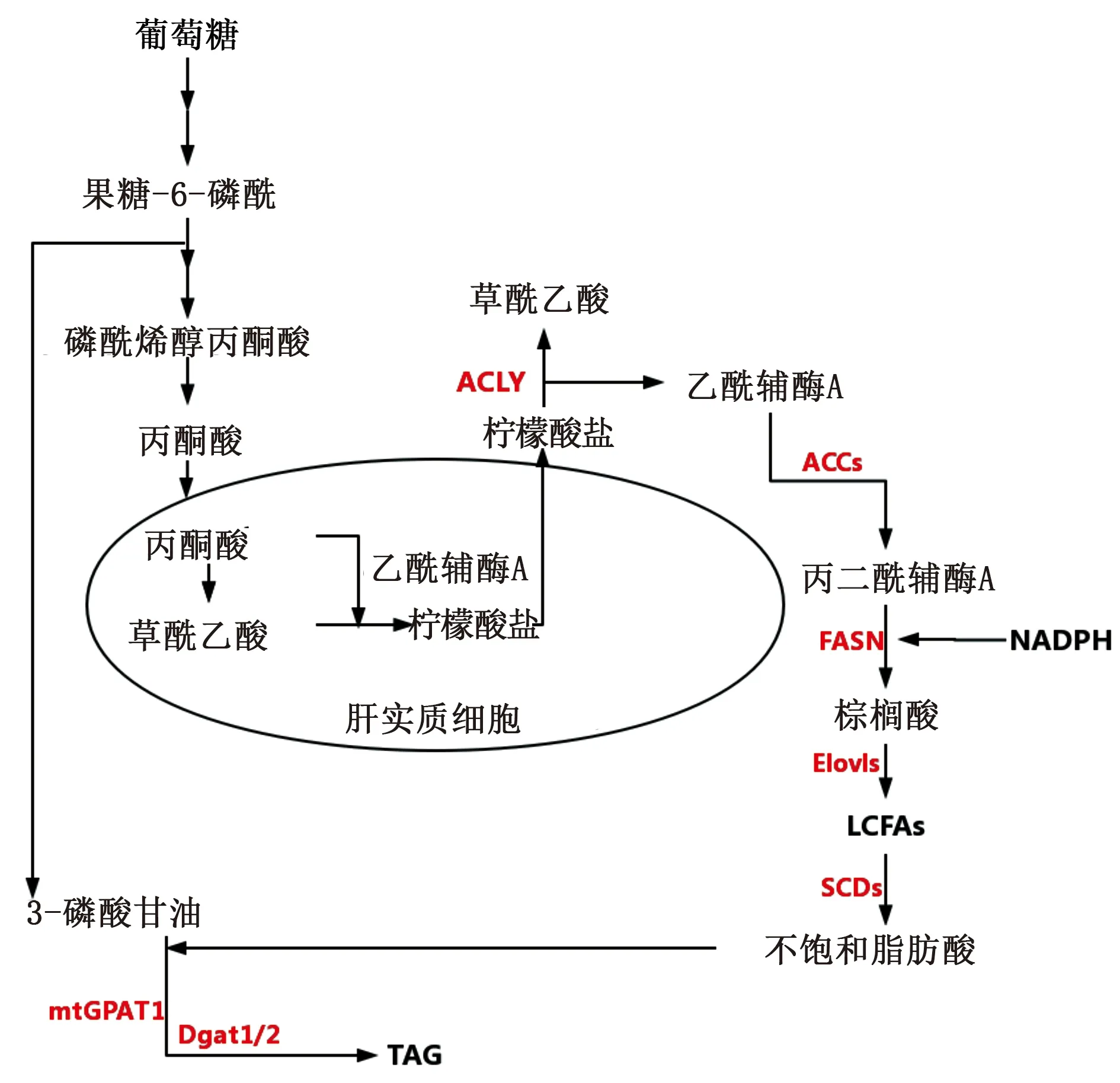
圖3 脂肪的從頭合成
從頭合成脂質是人體在進食時對能量進行儲存的方式,產生不飽和脂肪酸并和三磷酸甘油形成三酰基甘油儲存。從頭合成的脂質增多勢必會導致體內的TAG含量增多,誘導脂肪肝的產生。
關于參與脂質從頭合成過程相關酶的翻譯后修飾方式如下:
3.1 ATP-檸檬酸裂解酶(ACLY) ACLY是一種胞質酶,可催化檸檬酸生成乙酰輔酶A。若ACLY的水平提高,則會增加乙酰輔酶A的含量從而促進脂質的合成,導致了非酒精性脂肪肝等疾病的進一步發展[22]。
ACLY的翻譯后修飾包括了泛素化、乙酰化和磷酸化:①Cullin3(CUL3)是CUL3-RING泛素連接酶復合物的核心蛋白,CUL3可通過其銜接蛋白KLHL25(Kelch樣家族成員25)與ACLY相互作用,泛素化并降解細胞中的ACLY,通過對ACLY的負調節,抑制脂質合成[23];②ACLY在賴氨酸殘基540,546和554處被乙酰化。在高葡萄糖下,P300 /鈣結合蛋白(CBP)相關因子(P300/CBP associated factor,PCAF)乙酰轉移酶刺激這三個賴氨酸殘基的乙酰化,并通過阻斷其泛素化和降解來增加ACLY穩定性。相反,蛋白質脫乙酰基酶sirtuin 2(SIRT2)使ACLY去乙酰化并使其不穩定。其中,Lys-540為乙酰化和泛素化的競爭位點,兩種修飾起著相反的作用[24];③絲氨酸/蘇氨酸激酶1(AKT)可作用于ACLY的Ser-454,通過磷酸化激活ACLY,促進脂質的合成[25]。
3.2 乙酰輔酶A羧化酶1(ACC1) ACC1是ACC家族中的一種,可催化乙酰輔酶A的ATP依賴性羧化,這是脂肪酸生物合成的限速步驟。可見ACC1在脂肪的從頭合成中的重要性。抑制ACC1的活性就可減少內源性脂肪酸的生成,進而防止脂肪肝的進一步發展。
ACC1的翻譯后修飾的方式主要為磷酸化。
①AMP激活蛋白激酶(AMPK)AMPK可使 ACC1上的ser79磷酸化并抑制其活性,從而抑制了脂肪酸的合成[26];②TGFβ激活激酶(TAK)1 可介導ACC1磷酸化,TAK1可通過磷酸化AMPK的Thr-172位點使其激活,進而磷酸化ACC,降低ACC的活性[27];③蛋白磷酸酶2A異源三聚體(PP2A)可直接使AMPK的Thr-172去磷酸化而使AMPK活性降低,從而降低了ACC的磷酸化程度,提高了ACC的活性,促進脂質的合成積累[28];④核苷二磷酸激酶(NDPK)通過與AMPK競爭ATP的結合位點,調節AMPK的磷酸化程度,維持AMPK的活性,并調控AMPK體內的靶蛋白ACC1的磷酸化[29]。
3.3 脂肪酸合酶(Fasn) Fasn在內源脂肪酸的合成過程中,負責催化長鏈脂肪酸的合成,使丙二酰輔酶A與乙酰輔酶A聚合形成棕櫚酸,是脂肪酸合成過程中的關鍵酶。FASN活性的增強和減弱都會對脂質的從頭合成產生巨大的影響。許多實驗表明,從基因層面抑制FASN的表達可以減少體內脂肪酸的合成從而減輕脂肪肝的程度。另一方面,在翻譯后修飾的層面上,FASN受到磷酸化,乙酰化和泛素化的調節:①Fasn不同的磷酸化狀態可能不同地調節Fasn的活性,蘇氨酸的磷酸化可以抑制Fasn 的活性,而酪氨酸的磷酸化可以增強Fasn的活性。HRG(人表皮因子受體調節蛋白)可以激活HER2(人表皮生長因子受體2)酪氨酸激酶,從而導致誘導FASN的酪氨酸磷酸化,增強Fasn 的活性,而AMPK的磷酸化激活后可以磷酸化Fans中的蘇氨酸殘基,使得Fasn 的活性受到抑制,脂質生成減少[30];②酪氨酸磷酸酶Shp2作為連接分子可將泛素E3連接酶COP1(也稱為RFWD2)與FASN連接,從而加速FASN泛素化和降解,其中p38 MAP激酶可使COP1磷酸化,誘導COP1的核輸出,由細胞核至細胞質,參與FASN-Shp2-COP1復合物的形成[31]。而泛素特異性蛋白14(USP14)可阻斷FASN泛素化,增強FASN的穩定性[32];③賴氨酸乙酰轉移酶(KAT8)催化FASN乙酰化,然后促進FASN與 E3泛素連接酶TRIM21結合,導致隨后的多泛素化和蛋白酶體降解。相反,HDAC3(組蛋白脫乙酰基酶3)介導的FASN去乙酰化有助于提高FASN蛋白的水平,促進細胞中從頭脂肪酸的合成[33]。
3.4 硬脂酰輔酶A去飽和酶1(SCD1) SCD1可催化單不飽和脂肪酸(MUFA)的合成,主要是油酸和棕櫚油酸,它們在控制高碳水化合物飲食的體重增加中起重要作用[34]。實驗表明,SCD1在人體中的過度表達可能與高甘油三酯血癥(HTG),非酒精性脂肪肝,動脈粥樣硬化和糖尿病的病因有關,降低SCD1的活性可能是減少這些疾病的可行方法[35]。
SCD1的翻譯后修飾方式為:①表皮生長因子受體(EGFR)具有酪氨酸激酶的活性,可與SCD1在Y55結合并磷酸化SCD1,維持SCD1的穩定性,干擾SCD1的泛素化,增加MUFA(單不飽和脂肪酸)水平[36];②SCD1還可通過泛素-蛋白酶體依賴的ERAD途徑進行泛素化降解,SCD1末端含有66個殘基的富含PEST序列的N-末端區對于蛋白酶體依賴性十分重要,對于泛素化降解敏感[37]。
3.5 甘油-sn-3-磷酸酰基轉移酶1(mtGPAT1) mtGPAT1控制三酰基甘油(TAG)合成的第一步,對于慢性代謝紊亂如原發性非酒精性脂肪性肝病(NAFLD)至關重要[38]。抑制mtGPAT1的活性可稱為脂肪肝治療的靶點。
mtGPAT1主要受到磷酸化的修飾。
①酪氨酸激酶2(CK2)可磷酸化mtGPAT1的絲氨酸632、639位點,增強其活性[39];②增加蛋白激酶Cζ磷酸化和膜易位,可磷酸化mtF0F1-ATP酶β-亞基,降低其酶活性,從而抑制mtGPAT1活化[38];③AMPK可作用于mtGPAT1蛋白質的C末端結構域。AMPK磷酸化mtGPAT1可降低其活性[40]。
3.6 二酰基甘油酰基轉移酶1/2(DGAT1) DGAT1/2是一種完整的ER膜蛋白,可催化二酰基甘油和脂肪酰基CoA向三酰基甘油(TAG)的轉化。DGAT1突變導致其產物表達減少,導致先天性腹瀉疾病,這表明了DGAT1在脂質代謝中的重要作用[41]。
DGAT2主要修飾方式為泛素化,它有兩個跨膜結構域(TMD1和TMD2),可被蛋白酶體依賴性ERAD(ER相關的降解)途徑快速降解,其中,TMD1是DGAT2不穩定性的決定因素,其中的兩個賴氨酸殘基(K63和K66)首先被認為是泛素化的位點,被E3連接酶gp78(AMFR,autocrine motility factor receptor )識別并在ER處快速降解[42]。
4 脂肪的氧化
脂肪的氧化分解是機體功能的主要手段之一,脂質作為人體的主要儲能物質,主要以TG的形式儲存在人體的脂肪組織和VLDL中。當機體進行脂肪動員時,TG在脂肪水解酶ATGL、HSL等作用下分解成游離脂肪酸,經細胞攝取后在ACSL1的作用下活化,變為脂酰輔酶A,通過CPT1a進入線粒體,進行β-氧化,經過脫氫,加水,再脫氫,硫解四步后,分解為一分子的乙酰輔酶A和比原來少兩個碳的脂酰輔酶A,為機體提供不同長度的脂肪酸和能量[43](見圖4)。
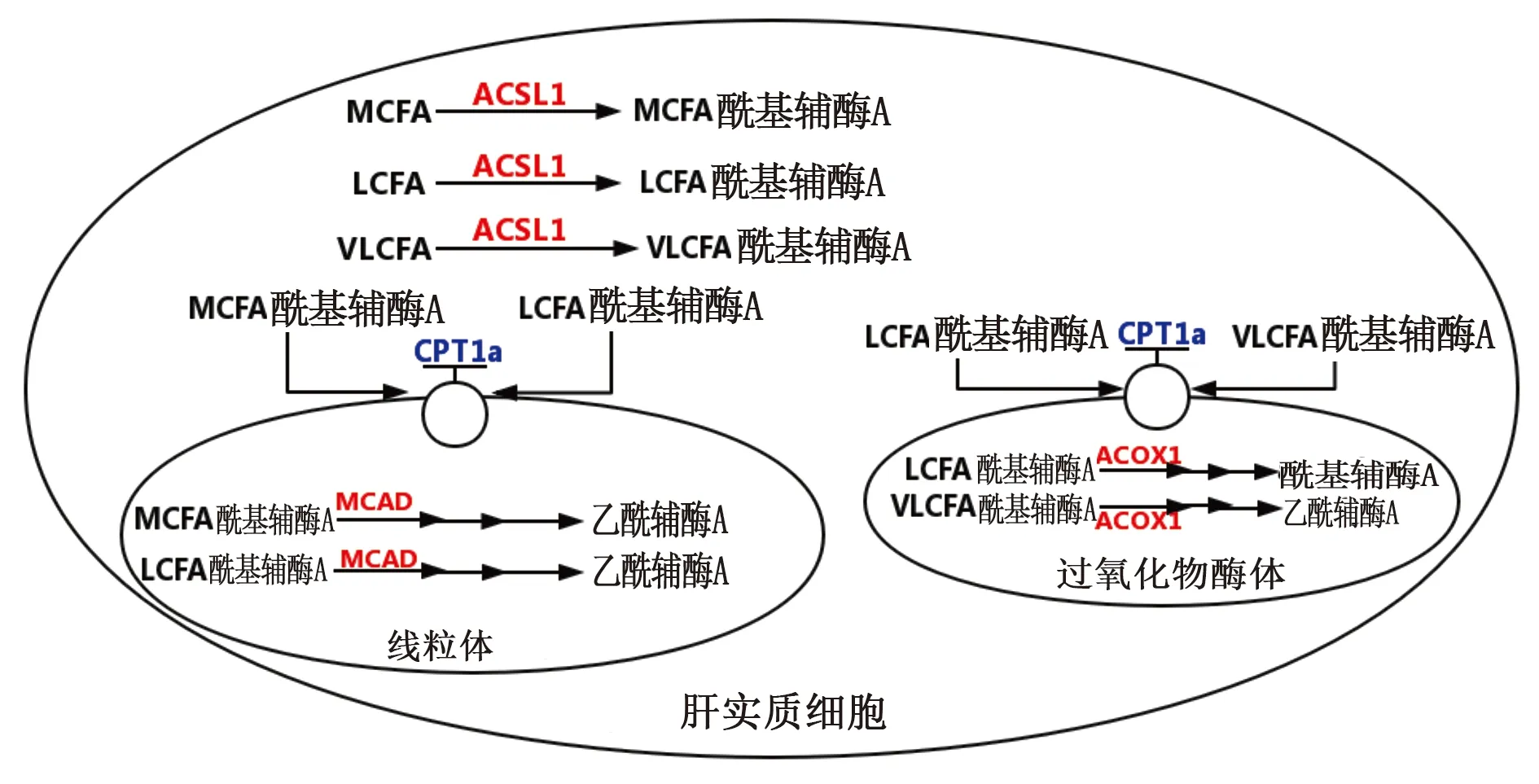
圖4 脂肪的氧化
若細胞對脂肪的氧化利用程度降低,脂肪動員減弱,就會導致脂質在細胞內部的過度沉積,誘導脂肪肝的產生。而影響氧化過程與參與反應的酶的活性密不可分。通過調節CPT1a等酶的活性,就能對脂肪的氧化進行調控,一定程度上抑制脂肪肝的進一步發展。
4.1 肉毒堿棕櫚酰轉移酶1a(CPT1a) CPT1a在脂質代謝中起作用,將LCFAs轉運到線粒體內進行氧化分解,釋放能量。而CPT1a缺乏癥是一種罕見的長鏈脂肪酸氧化常染色體隱性遺傳病。在禁食或生病期間,患者有發生低血糖和肝功能衰竭的風險[44],可見CPT1a在脂肪氧化中的重要作用。
CPT1a的活性可受到丙二酰輔酶A的抑制,而AMPK可磷酸化ACC,使ACC活性降低,降低丙二酰輔酶A的水平,從而提高了CPT1a的活性[45]。
4.2 中鏈酰基輔酶A脫氫酶(MCAD) MCAD是一種對脂肪酸代謝至關重要的線粒體基質蛋白,在脂質的氧化過程中起重要作用。若MCAD的含量與活性改變,會使得機體的能量代謝產生紊亂。MCAD缺乏癥是最常見的脂肪酸氧化病,患者可能出現急性高氨血癥、低血糖、腦病和肝大,主要是在長時間的并發感染后。也可能發生與心跳障礙有關的猝死[46]。
MCAD主要通過磷酸化進行翻譯后修飾。PTEN誘導的假定激酶1(PINK1)是一種線粒體靶向激酶,PINK1是絲氨酸/蘇氨酸激酶,可磷酸化MCAD的S347,增強其活性從而促進MCAD對中鏈脂肪酸的氧化[47]。
4.3 長鏈酰基輔酶A脫氫酶(LCAD) LCAD是參與脂肪酸氧化的關鍵酶,它與MCAD類似,對于脂肪酸的氧化分解有重要意義。有實驗證明LCAD缺乏的小鼠能量代謝效率降低,對葡萄糖的依賴性提高,易感與生物饑餓的流感最終消瘦死亡[48]。可見LCAD在能量代謝中的重要性,人體缺失LCAD可能將導致機體供能不足而影響功能。
SIRT3是一種線粒體NAD+依賴性脫乙酰酶,可通過使LCAD的賴氨酸42殘基去乙酰化,提高LCAD的活性,減少脂質的積累[49]。
4.4 酰基輔酶A氧化酶1(ACOX1) ACOX1是脂肪酸β-氧化中的第一種限速酶,是β-氧化不可或缺的部分。但它參與的不是線粒體的β-氧化,而是在過氧化物酶體中的不同于線粒體的β氧化,底物主要包括了極長鏈脂肪酸、長鏈脂肪酸、2-甲基支鏈脂肪酸等。其β-氧化的過程包括了氧化、水化、脫氫和硫解,可合成體內所必需的生理活性物質,但氧化不徹底,通常是進行β-氧化循環而縮短碳鏈[50]。抑制了ACOX1的活性將減少極長鏈脂肪酸等進行氧化,導致過多的脂肪積累,誘導脂肪肝的產生。
脫乙酰酶sirtuin 5(SIRT5)介導的賴氨酸脫琥珀酸化可通過使ACOX1中的賴氨酸脫琥珀酸乙酰化,損害其活性二聚體引起ACOX1的酶抑制,防止出現脂質過氧化帶來的機體氧化損傷[51]。
總之,脂肪肝的發生和發展是一個復雜的過程,受到多種途徑和機制的共同影響,而脂質代謝的紊亂是脂肪肝發生的重要原因,了解脂質的轉運,合成及氧化過程,可為脂肪肝的研究和治療提供十分有用的線索。本文分別總結了在脂質代謝的3個不同步驟——轉運攝取,從頭合成和脂肪氧化中起作用的酶和蛋白質的翻譯后修飾的方式及其功能,可能對于脂肪肝的研究有一定的幫助。當然,這些蛋白質的翻譯后修飾是否在脂肪肝中起變化還需要試驗的證明,同時,這些蛋白質在基因水平的變化也是研究脂肪肝的過程中需要關注的地方,對于脂肪肝的研究,還有許多事情等著我們。
