耐鹽菌發酵對咸鴨蛋蛋清脫鹽及抗氧化活性的影響
2020-01-19 03:15:16陳遠哲黃熙鶯魯安娜尉鑫欣王柳雄龔金炎肖功年
食品與生物技術學報 2019年11期
關鍵詞:質量
陳遠哲, 黃熙鶯, 魯安娜, 尉鑫欣, 王柳雄, 龔金炎, 肖功年*
(1. 浙江科技學院 省農產品化學與生物加工技術重點實驗室,浙江 杭州310023;2. 嘉興市家家樂食品有限責任公司,浙江 嘉興314018)
我國屬于禽蛋生產大國, 鴨蛋產量居世界首位。 浙江省粽子產業發達,然而每年因生產咸蛋黃粽而遺留下數萬噸咸蛋清, 由于其高鹽質量分數(7%~12%)以及黏度高,始終無法得到有效地利用。鴨蛋清蛋白是一種優質的蛋白質資源,其氨基酸組成與雞蛋清相似[1]。
目前對咸鴨蛋清的處理除直接用于高品質面條[2],重組魚粒[3-4]外,還將其酶解脫鹽,通過酶解產生的小肽類物質大大提高其附加值。 其中最關鍵的一步便是脫鹽,然而常規的脫鹽手法如超濾[5]、電滲析等方法,存在堵塞和黏附等問題,同時由于咸蛋清營養價值豐富,易于微生物繁殖,一旦污染導致濾膜難清洗、再生困難等問題。 先酶解[6],雖然在一定程度上能降低咸鴨蛋蛋清黏度,緩解了膜技術脫鹽過程存在的過濾膜易堵塞的問題,但在高濃度的鹽環境中,酶活性會降低或受到抑制,酶解效率降低,酶解咸蛋清的酶及酶解參數,更是難以選擇和調控[7]。
作者從腌制食品中篩選出耐鹽菌株Staphylococcus equorum,接種于咸鴨蛋清中,在最適條件下發酵。 發酵法的優勢是將微生物產酶和酶水解相結合,簡化生產工藝,降低成本,極大得提高咸鴨蛋蛋清的附加值,為咸鴨蛋蛋清的再生利用提供一條新思路。
1 材料與方法
1.1 材料與儀器
金華火腿、咸鴨蛋蛋清:市售;10%含鹽質量分數的MSA 培養基、牛血清蛋白、VC(Trolox)、乙二胺四乙酸二鈉(分析純)、抗壞血酸(分析純)、二苯代苦味酰基自由基(DPPH·)、鐵氰化鉀(分析純)、三氯乙酸(分析純)、氯化鐵(分析純)、菲啰嗪、氯化亞鐵(分析純)、冰乙酸(分析純)、硫代巴比妥酸(分析純)、軟磷脂(分析純)、氫氧化鈉(分析純)。
DV-S 型數顯粘度計:美國brookfield;UV-5200PC型紫外分光光譜:上海元析儀器有限公司;TG16K-Ⅱ型離心機:上海趙迪生物科有限公司;UDK159 凱式定氮儀:VELP 公司;DK20 消化爐:VELP 公司;真空冷凍干燥機;Millipore 超濾離心管。
1.2 實驗方法
1.2.1 不同發酵時間咸鴨蛋蛋清粘度的變化參照葉青松[8]等人的方法,取適量待測樣品,置于100 mL 的燒杯中,選用s61 號轉子,轉速100 r/min,測定黏度值。
1.2.2 咸蛋清發酵產物制備分別取適量鴨蛋清和沉淀物加去離子水振蕩30 min, 離心取上清液,0.22 um 膜過濾后, 分別用10 000 和3 000 的超濾膜離心截留分離,保留濃縮液,分別標記為組分Ⅰ、Ⅱ、Ⅲ(空白組),發酵組Ⅰ、Ⅱ、Ⅲ,凍干備用。
1.3 測試方法
1.3.1 鹽質量分數及脫鹽率測定參照文獻[9]的方法。
1.3.2 蛋白質質量分數測定可溶性蛋白質質量分數和粗蛋白質質量分數,分別采用考馬斯亮藍法[10]和凱氏定氮法[11]測定。
1.3.3 DPPH·清除能力測定參照文獻[12]的方法。
1.3.4 Fe2+螯合能力的測定參照文獻[13]的方法。
1.3.5 抑制脂過氧化能力的測定參照文獻[14]的方法。
1.3.6 還原力測定參照文獻[15]的方法。
2 結果與討論
2.1 咸鴨蛋蛋清發酵后理化指標的測定
2.1.1 咸鴨蛋蛋清發酵過程黏度的變化咸鴨蛋去掉蛋黃后蛋清一般比較粘稠,據葉青松等人的報道,咸鴨蛋蛋清不經處理,黏度在80 mPa·s 左右,黏度是表征咸蛋清膜法脫鹽效率的重要特征之一[16],Staphylococcus equorum不同發酵時間對黏度的影響見圖1。
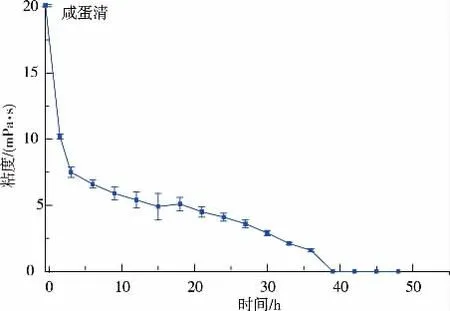
圖1 咸蛋清不同發酵時間粘度的變化Fig. 1 Change of viscosity of different fermentation
鴨蛋經過腌制,蛋清蛋白逐漸水樣化,但仍具有一定的黏度,從圖1 中得知,0 h 咸鴨蛋清的黏度在20 mPa·s 左右,無論是對膜脫鹽還是膜過濾都會造成膜孔堵塞等嚴重影響[17]。 接入Staphylococcus equorum菌懸液,發酵初期,在微生物的作用下黏度迅速下降,隨著發酵時間的增長,逐漸趨于平緩,但黏度仍不斷的降低,最終達到零。
2.1.2 發酵不同時間pH 的變化, 上清液及離心沉淀中的鹽質量分數發酵過程中pH 的變化見圖2。 本次實驗所用的咸鴨蛋蛋清鹽質量分數為10%,每隔12 小時取一次樣測pH 值,離心后對上清液及沉淀物的鹽質量分數進行測定。 由圖3 可知,沉淀物中鹽質量分數大約在3%左右。 咸蛋清發酵過程中,耐鹽菌分泌的蛋白酶破壞蛋白質的結構[18],蛋清蛋白與水形成的膠狀結構被破壞,水分析出帶走部分鹽分;隨著pH 的降低,部分蛋白質及肽類物質,達到其等電點并形成沉淀[19],此時發酵液已明顯分層,通過離心取沉淀物,大部分鹽仍留在上清液中,從而達到脫鹽的目的。3%左右的鹽質量分數與新鮮咸蛋清相比,脫鹽率可達70%,且擠出多余的水分后沉淀物中的鹽質量分數會進一步降低。 沉淀物凍干粉中的鹽質量分數只有15%,也已遠遠低于咸蛋清凍干粉中38%的鹽質量分數,見圖4。
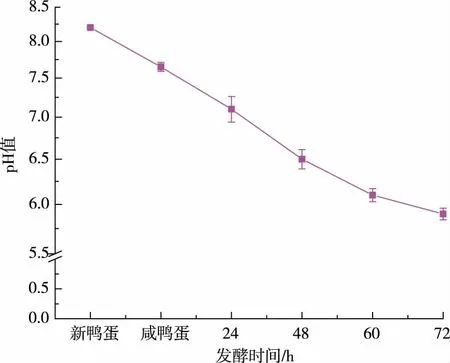
圖2 發酵不同時間pH 的變化Fig. 2 pH of different fermentation
2.1.3 離心沉淀物凍干粗蛋白質質量分數由圖5可知,0 小時代表新鮮咸鴨蛋蛋清,由于該耐鹽菌株在生長過程中產酸,沉淀物中蛋白質來源大致有三部分組成,菌體、遇酸沉淀的蛋白質以及部分不可溶的蛋清蛋白。發酵36 h 的沉淀物粗蛋白質質量分數高于凍干咸蛋清粉,鹽質量分數大大低于凍干咸蛋清,是一種很好的高蛋白、低含鹽質量分數的飼料添加成分。 隨著發酵時間的延長,微生物活動消耗,蛋白質質量分數逐漸降低。
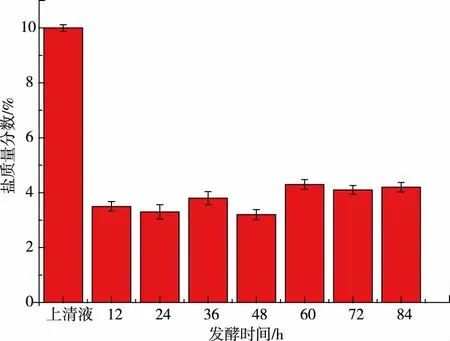
圖3 發酵不同時間上清液及離心沉淀中的鹽質量分數Fig. 3 Salt content of different fermentation of the supernatant and precipitate
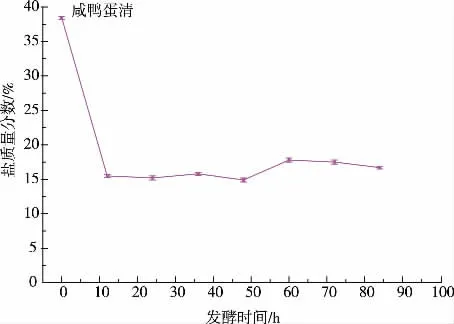
圖4 發酵不同時間沉淀物凍干粉鹽質量分數Fig. 4 Salt content of differentfermentation of the freezedried precipitate
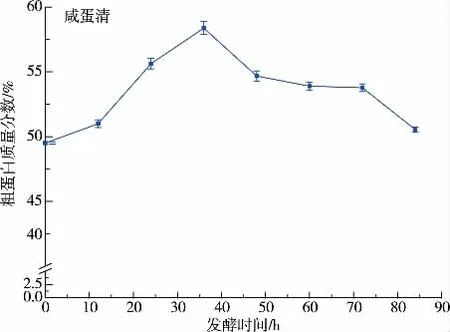
圖5 發酵不同時間沉淀物凍干粗蛋白質量分數Fig. 5 Protein content ofdifferent fermentation of the freeze-dried precipitate
2.2 咸蛋清發酵產物抗氧化活性的測定
2.2.1 蛋白質質量濃度測定的標準曲線如圖6所示,表明在0~0.1 mg 范圍內牛血清蛋白的質量濃度與吸光度呈較好的線性關系, 線性回歸方程為Y=6.982 6X+,0.009 17,R2=0.997 0。
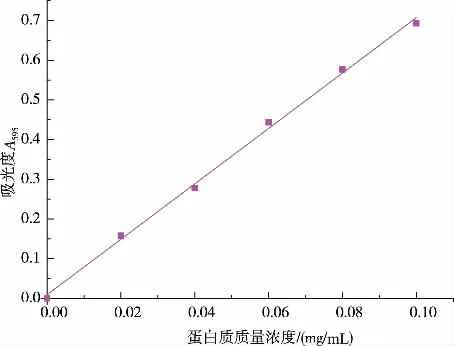
圖6 牛血清蛋白質標準曲線Fig. 6 Standard curve of bovine serum albumin solution
2.2.2 發酵產物DPPH·清除能力測定二苯代苦味酰基自由基(DPPH·)是一種穩定的自由基,在波長517 nm 處有最大吸收峰, 當體系中有強抗氧化劑存在時,孤對電子會被配對,吸光值下降,其褪色程度與結合的電子數有定量關系[20],見圖7。
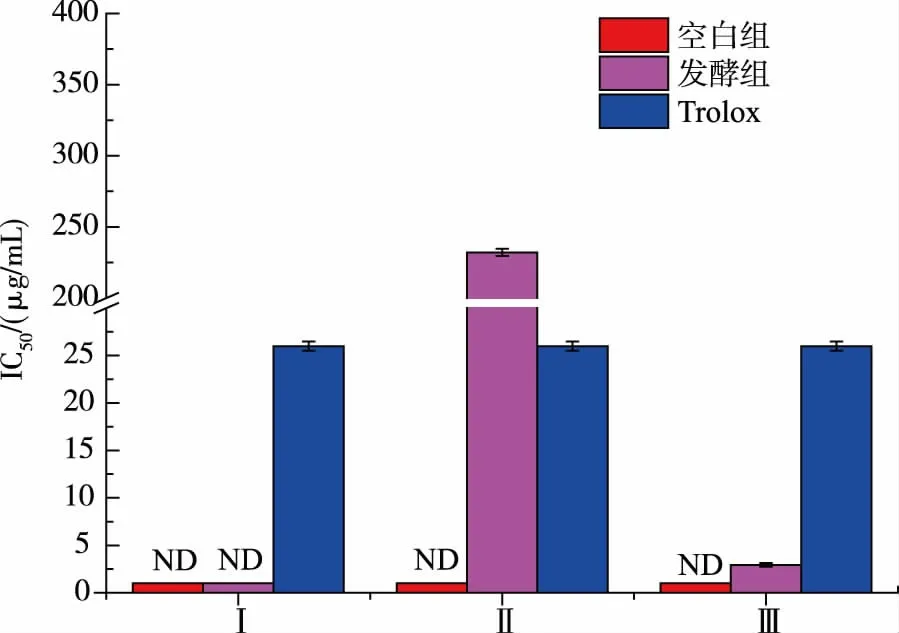
圖7 發酵產物體外DPPH·自由基清除活性Fig. 7 Scavenging activity of different fermentation samples to DPPH·in vitro
比較圖7 中空白組和發酵組間發現,發酵組組分Ⅲ的IC50值遠低于對照組,表現出較強的抗氧化活性, 有研究發現一些特定氨基酸 (如Glu、 Asp、Lys、Leu 和Ala) 的存在能夠增強肽的抗氧化性。Lys 等因其側鏈有氨基或羧基, 而具有清除自由基和螯合金屬離子的能力[21-22]。 空白組由于蛋清蛋白相對分子質量均在10 000 以上,故超濾離心后得不到組分Ⅱ、Ⅲ,檢測不到抗氧化活性。 發酵組組分Ⅰ檢測不到活性的原因,猜測是因為具有抗氧化活性的小肽含量比較少,且復雜的環境成分對實驗有一定的干擾。 由此可得出耐鹽菌發酵后能增強咸蛋清蛋白的抗氧化活性。
2.2.3 發酵產物Fe2+螯合能力的測定比較圖8 中空白組和發酵組發現,其還原力與相對分子質量大小有一定關系。相對分子質量越小,還原力越強。發酵組組分Ⅲ的IC50遠小于對照組,具有極強的抗氧化活性,較空白組而言,是從無到有的突破;發酵組組分Ⅱ的IC50雖然大于對照組, 但與空白組相比較,仍具有較強的抗氧化活性。 空白組的三個組分均檢測不到抗氧化活性,由此可得出,耐鹽菌發酵有助于增強咸蛋清蛋白的抗氧化活性。 有研究表明[23-24],Fe2+螯合能力不僅與樣品中雜環化合物的含量也有較大的關聯,與樣品中的大分子物質也有很大的關聯。
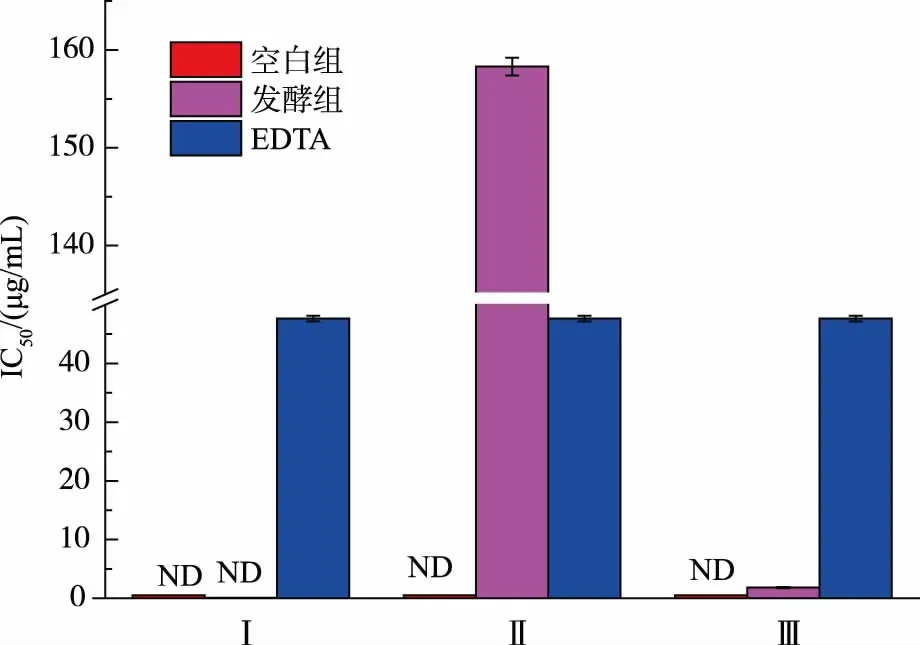
圖8 發酵產物體外Fe2+螯合能力Fig. 8 Fe2 + chelating ability of different fermentation samples in vitro
2.2.4 發酵產物抗脂質過氧化能力的測定該方法利用多不飽和脂肪酸在氧化過程中,產生的丙二醛和硫代巴比妥酸反應生成紅色物質, 該物質在532 nm 處有強吸收峰。抗氧化劑能抑制多不飽和脂肪酸的自動氧化,丙二醛的生成量隨之降低,紅色變淡甚至消失,因此可以通過測定反應溶液的吸光值來檢測產物的抗氧化效果[25],見圖9。
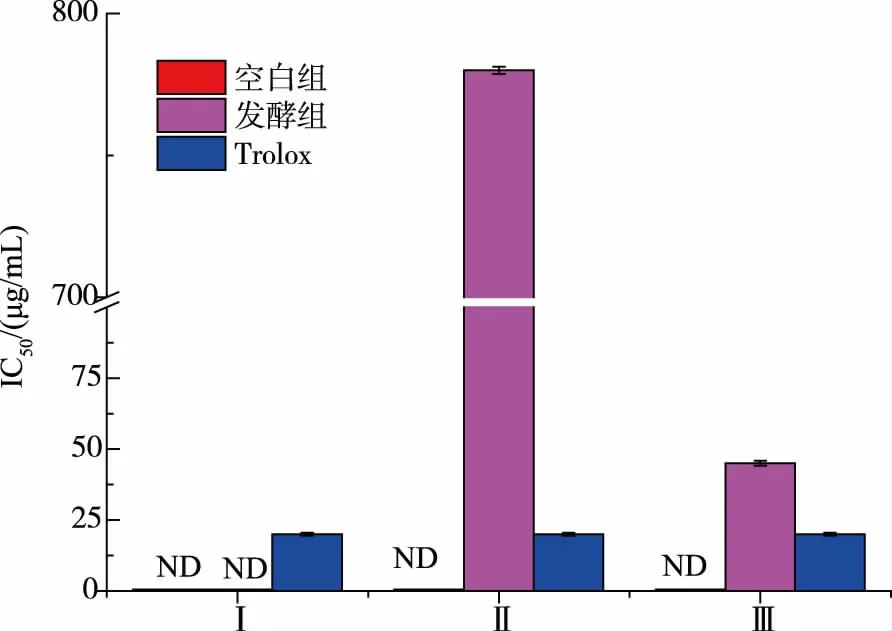
圖9 發酵產物體外抑制脂質過氧化能力Fig. 9 Inhibiting lipid peroxidation ability of different fermentation samples in vitro
從圖9 可知, 在微生物分泌的蛋白酶的作用下,咸鴨蛋蛋清具有抗脂質過氧化能力的基團逐漸暴露出來,表現出了一定的抗氧化活性[26]。但從抑制脂質過氧化能力來看,發酵組組分Ⅱ、Ⅲ的IC50值相較于對照組來說并不是很優秀,對空白組而言卻有質的突破。 發酵組組分Ⅰ檢測不到活性的原因,猜測是因為具有抗氧化活性的小肽含量相對較少,且復雜的環境成分對實驗有一定的干擾。 由此可得出,耐鹽菌發酵有助于增強咸蛋清蛋白的抑制脂質過氧化能力。
2.2.5 發酵產物還原力測定具有抗氧化能力的物質提供的電子, 能使Fe3+還原成Fe2+,K3Fe(CN)6在還原劑的作用下被還原成K4Fe(CN)6,與Fe3+反應能形成普魯士藍,其在700 nm 附近有強吸收峰[27]。
還原力是基于加入發酵離心產物 (還原性物質)后,體系中Fe3+轉化為Fe2+來檢測的,吸光值越大樣品的還原力越強。 如圖10 所示,VC 當量越大表明樣品還原力越強,比較發酵組組分Ⅱ、Ⅲ可知,其還原力與相對分子質量大小呈負相關,相對分子質量在3 000 以下, 在蛋白酶的作用下一些極性或帶電的氨基酸側鏈較多地暴露出來[28],從而增加了還原能力。 比較發酵組的組分Ⅱ、Ⅲ,其還原力隨相對分子質量的減小而增強,這與Guillen G 等人[29]的研究結果一致。 空白組的三個組分均檢測不出還原能力。 由此可得出,耐鹽菌發酵后能增強咸蛋清蛋白的抗氧化活性。
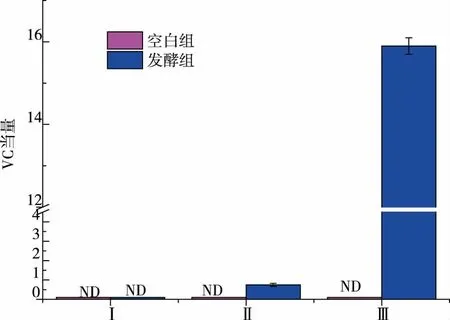
圖10 發酵產物體外還原力Fig. 10 Reducing ability of different fermentation samples in vitro
3 結 語
從金華火腿中篩選得到的耐鹽菌,37 ℃振蕩發酵48 h,6 000 r/min 離心取沉淀物, 蛋白質質量分數約為12%,與新鮮鴨蛋清無異,但鹽質量分數僅剩3%左右, 脫鹽率能達到70%, 相較于咸鴨蛋清10%的含鹽質量分數大大降低;凍干后,鹽質量分數15%~18%,大大低于咸蛋清38%;粗蛋白質量分數55%~58%,略高于咸蛋清49%。
沉淀物加水振蕩復溶, 取上清液分別經過10 000 和3 000 的超濾膜離心分離, 得到小于3 000 及3 000~10 000 的物質,通過四個體外抗氧化活性體系共同得出, 咸鴨蛋蛋清在發酵過程中,耐鹽菌產生的蛋白酶能在較高鹽質量分數的環境中有效地降解蛋清蛋白, 經過3 000 的超濾膜離心后得到小于3 000 的小肽,具有極強的抗氧化活性。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
