桑黃原生質(zhì)體的制備與再生*
2020-01-15 03:20:10孫婷婷王世新王旭彤劉增才馬伊莎
中國(guó)食用菌 2019年12期
孫婷婷,王世新,王旭彤,劉增才,馬伊莎,鄒 莉**
(1.哈爾濱學(xué)院 食品工程學(xué)院,黑龍江 哈爾濱 150086;2.東北林業(yè)大學(xué) 林學(xué)院,黑龍江 哈爾濱 150040)
桑黃(Phellinus igniarius)是一種珍稀的藥用真菌,含有多種活性成分,如多糖、三萜、黃酮等[1-5],在抗腫瘤、保肝護(hù)肝、降血糖、治療炎癥、抗衰老及增強(qiáng)機(jī)體免疫力等方面具有很高的藥用價(jià)值[6-11]。目前在生物抗癌領(lǐng)域中,桑黃已被公認(rèn)為防癌、抗癌效果最好的高等真菌之一。在韓國(guó)和日本的醫(yī)藥行業(yè)中,已廣泛使用桑黃治療及預(yù)防各類(lèi)癌癥的發(fā)生。
桑黃屬于多年生真菌,生長(zhǎng)周期長(zhǎng),生物學(xué)特性復(fù)雜[12],加之其人工栽培尚不成規(guī)模,因此導(dǎo)致市售桑黃子實(shí)體的價(jià)格昂貴、供不應(yīng)求。此外,液體發(fā)酵生產(chǎn)桑黃菌絲過(guò)程中仍存在產(chǎn)量和質(zhì)量不穩(wěn)定、菌種易退化等問(wèn)題,因此選育出性狀優(yōu)良、高產(chǎn)的桑黃菌株對(duì)其進(jìn)一步開(kāi)發(fā)利用具有重要意義[13]。桑黃的雜交育種工作由于缺少穩(wěn)定遺傳的分子標(biāo)記且遺傳背景復(fù)雜而進(jìn)展緩慢。目前,作者所在課題組完成了桑黃的轉(zhuǎn)錄組測(cè)序,并對(duì)其進(jìn)行了深入分析,為桑黃的分子遺傳學(xué)和功能基因組研究提供數(shù)據(jù)支持[14]。
制備優(yōu)質(zhì)高產(chǎn)的原生質(zhì)體不僅是分子遺傳學(xué)研究的關(guān)鍵步驟,同時(shí)也是菌株誘變、融合育種、基因轉(zhuǎn)化等操作的基礎(chǔ)和前提,因此建立穩(wěn)定高產(chǎn)的原生質(zhì)體制備技術(shù)尤為重要[15]。近年來(lái),桑黃原生質(zhì)體制備和再生優(yōu)化體系的建立鮮有報(bào)道。本研究采用桑黃菌株DL101 作為試驗(yàn)材料,詳細(xì)、系統(tǒng)地探討了桑黃原生質(zhì)體的制備與再生條件,優(yōu)化各試驗(yàn)因素,盡可能獲得更多的優(yōu)良原生質(zhì)體,旨在為桑黃后續(xù)的分子遺傳轉(zhuǎn)化、基因編輯及菌種改良打下基礎(chǔ)。
1 材料與方法
1.1 供試菌株及試劑
供試菌株桑黃DL101 保存于東北林業(yè)大學(xué)林學(xué)院森林保護(hù)試驗(yàn)室。溶壁酶購(gòu)自廣東微生物研究所;崩潰酶購(gòu)自Sigma 公司;葡萄糖、甘露醇、無(wú)水硫酸鎂、氯化鈉、瓊脂等均為國(guó)產(chǎn)分析純。
1.2 培養(yǎng)基
1.2.1 基礎(chǔ)培養(yǎng)基
PD 培養(yǎng)基:每升培養(yǎng)基中含有馬鈴薯(去皮)200 g、葡萄糖20 g。
PDA 培養(yǎng)基: 每升培養(yǎng)基中含有馬鈴薯(去皮)200 g、葡萄糖20 g、瓊脂粉15 g~18 g。
1.2.2 再生培養(yǎng)基
PDA+甘露醇再生培養(yǎng)基:在上述基礎(chǔ)培養(yǎng)基PDA 中加入甘露醇109.30 g·L-1。
PDA+硫酸鎂再生培養(yǎng)基:在上述基礎(chǔ)培養(yǎng)基PDA 中加入無(wú)水硫酸鎂72.22 g·L-1。
PDA+氯化鈉再生培養(yǎng)基:在上述基礎(chǔ)培養(yǎng)基PDA 中加入氯化鈉35.06 g·L-1。
1.3 菌絲培養(yǎng)
將培養(yǎng)8 d 的桑黃菌絲PDA 平板,用打孔器打出菌餅后接種于200 mL PD 培養(yǎng)基中,25℃、180 r·min-1振蕩培養(yǎng)5 d,用無(wú)菌紗布過(guò)濾,并用無(wú)菌水沖洗2 次~3 次,收集菌絲,置于新鮮PD 培養(yǎng)基中,4℃冰箱保存?zhèn)溆谩?/p>
1.4 原生質(zhì)體的制備
稱取上述適量濕菌絲于2 mL 離心管中,4 000 r·min-1離心5 min,棄去上清液,再用0.6 mol·L-1甘露醇滲透壓穩(wěn)定劑溶液洗滌、離心2 次,條件同上,獲得干凈的菌絲0.1 g,然后加入 1 mL 酶液,26℃~34℃水浴酶解2.0 h ~4.0 h,每隔0.5 h 對(duì)酶解液顛倒混勻數(shù)次。吸取10 μL 酶解液,血球計(jì)數(shù)板計(jì)數(shù),鏡檢破壁情況。
1.4.1 單因素試驗(yàn)
按照上述方法, 首先分別考察不同的酶濃度(1.0%溶壁酶、1.5%溶壁酶、2.0%溶壁酶、2.5%溶壁酶、3%溶壁酶)、不同的酶系統(tǒng)(0.5%崩潰酶+溶壁酶、1.0%崩潰酶+溶壁酶)對(duì)原生質(zhì)體得率的影響。然后在確定酶系統(tǒng)的基礎(chǔ)上再分別考察不同的酶解溫度(26℃、28℃、30℃、32℃、34℃)、不同的酶解時(shí)間(2.0 h、2.5 h、3.0 h、3.5 h、4.0 h)對(duì)原生質(zhì)體得率的影響。
1.4.2 響應(yīng)面優(yōu)化試驗(yàn)
采用響應(yīng)面法對(duì)桑黃原生質(zhì)體的制備條件進(jìn)行優(yōu)化。根據(jù)單因素試驗(yàn)結(jié)果, 利用Box-Behnken 設(shè)計(jì)試驗(yàn),自變量選取酶解溫度(A),分別設(shè)為28℃、30℃、32℃;酶解時(shí)間(B),分別設(shè)為2.5 h、3.0 h、3.5 h;溶壁酶的濃度(C),分別設(shè)為1.5%、2.0%、2.5%,因變量即響應(yīng)值選取原生質(zhì)體的得率(Y)建立回歸方程,Box-Behnken 設(shè)計(jì)方案及原生質(zhì)體的產(chǎn)量結(jié)果,見(jiàn)表1,試驗(yàn)點(diǎn)共計(jì) 17 個(gè),其中分析點(diǎn)12 個(gè),零點(diǎn)5 個(gè)。
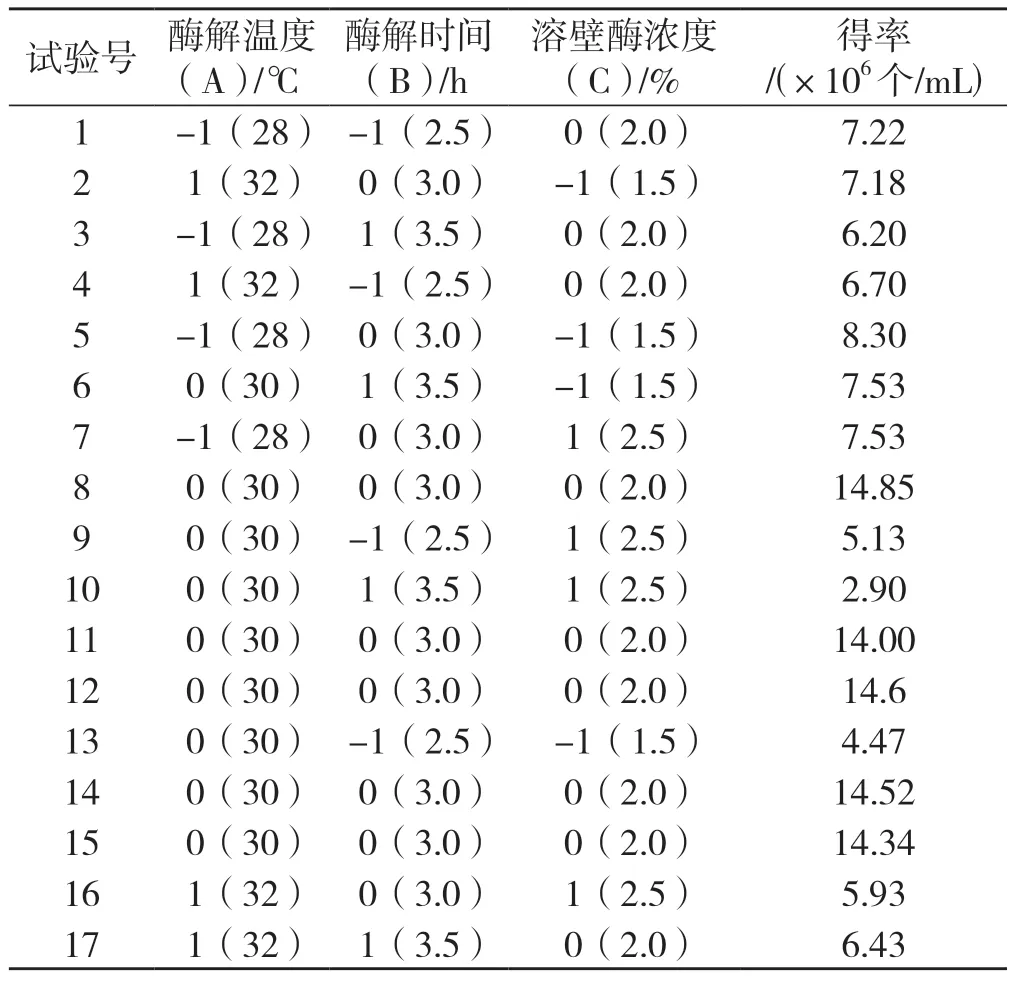
表1 Box-Behnken 設(shè)計(jì)方案及結(jié)果Tab.1 The design matrix and results of Box-Behnken experiment
1.5 原生質(zhì)體的再生
將酶解液用無(wú)菌的G3 砂芯漏斗過(guò)濾,除去殘留菌絲,4 000 r·min-1離心10 min,去上清液,用滲透壓穩(wěn)定劑洗滌2 次,將原生質(zhì)體用滲透壓穩(wěn)定劑制成懸浮液,在顯微鏡下用血球計(jì)數(shù)板計(jì)數(shù)。然后將得到的原生質(zhì)體懸浮液稀釋至105個(gè)/mL,取100 μL 分別涂布于PDA+甘露醇再生培養(yǎng)基、PDA+硫酸鎂再生培養(yǎng)基、PDA+氯化鈉再生培養(yǎng)基和PDA 培養(yǎng)基(對(duì)照),置于25℃條件下再生培養(yǎng),菌落計(jì)數(shù)。
再生率的計(jì)算:將制備的原生質(zhì)體分α 組、β組進(jìn)行再生對(duì)比試驗(yàn),α 組用滲透壓穩(wěn)定劑配制再生培養(yǎng)基,β 組用純水配制再生培養(yǎng)基進(jìn)行再生,每組設(shè)3 次重復(fù), 待菌落長(zhǎng)出后分別計(jì)數(shù), 取平均值,按下述方法計(jì)算再生率。再生率(P,%)公式為:
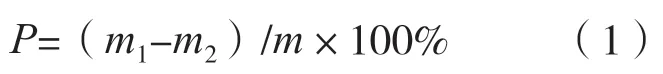
式中:m1表示α 組再生菌落數(shù);m2表示β 組再菌落數(shù);m表示原生質(zhì)體總數(shù)。
2 結(jié)果與分析
2.1 單因素試驗(yàn)結(jié)果
2.1.1 酶濃度及酶系統(tǒng)對(duì)桑黃原生質(zhì)體制備的影響
溶壁酶作用于菌絲體的細(xì)胞壁,將致密、完整的細(xì)胞壁結(jié)構(gòu)瓦解,釋放出一個(gè)個(gè)球狀的原生質(zhì)體,如圖1 所示。不同濃度的溶壁酶作用效果不同,見(jiàn)圖2。
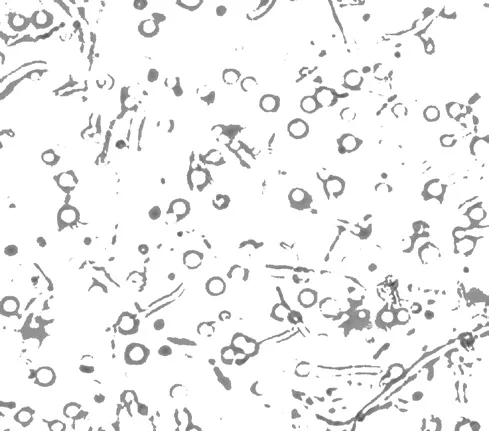
圖1 桑黃原生質(zhì)體(10×40)Fig.1 Protoplast of Phellinus igniarius(10×40)
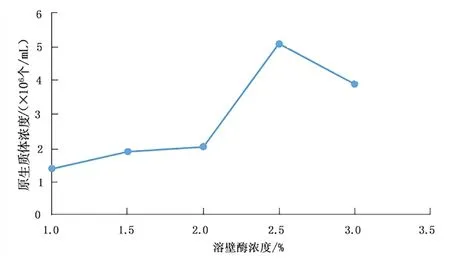
圖2 溶壁酶濃度對(duì)桑黃原生質(zhì)體制備的影響Fig.2 Effects of lywallzyme concentration on protoplast preparation of Phellinus igniarius
由圖1、圖2 可以看出,在一定范圍內(nèi),溶壁酶的濃度越高,釋放出的原生質(zhì)體越多,超過(guò)一定范圍,溶壁酶的濃度增加,原生質(zhì)體的數(shù)目減少。原因可能是溶壁酶對(duì)于原生質(zhì)體有一定的傷害作用,當(dāng)溶壁酶的濃度在2.5%以內(nèi)時(shí),隨著濃度增加,原生質(zhì)體釋放的速率大于原生質(zhì)體被破壞的速率,故原生質(zhì)體濃度不斷增加,超過(guò)這個(gè)范圍,原生質(zhì)體被破壞的速率大于原生質(zhì)體釋放的速率,故原生質(zhì)體濃度降低。當(dāng)溶壁酶濃度為2.5%時(shí),原生質(zhì)體得率最高,為5.1×106個(gè)/mL。
許多研究表明制備原生質(zhì)體時(shí),使用復(fù)合酶液的效果大于單一酶液。本試驗(yàn)采用不同濃度的崩潰酶和溶壁酶的組合,研究其對(duì)桑黃原生質(zhì)體制備的影響,結(jié)果如圖3 所示。
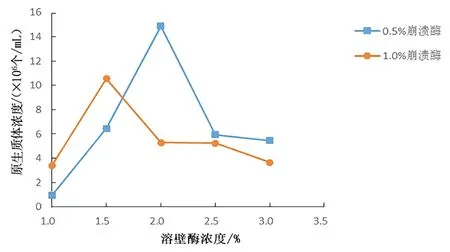
圖 3 不同濃度的崩潰酶和溶壁酶的組合對(duì)桑黃原生質(zhì)體制備的影響Fig.3 Effects of different combinations of driselase and lywallzyme on protoplast preparation of Phellinus igniarius
結(jié)合圖2,由圖3 可以發(fā)現(xiàn)崩潰酶與溶壁酶的復(fù)合酶液的作用效果確實(shí)優(yōu)于單一的溶壁酶,使用復(fù)合酶液,桑黃原生質(zhì)體得率最高可達(dá)到107個(gè)/mL,遠(yuǎn)大于單獨(dú)使用溶壁酶的作用效果。從圖3 中可以發(fā)現(xiàn),1.0%崩潰酶與低濃度的溶壁酶組合的效果優(yōu)于0.5%崩潰酶與低濃度的溶壁酶組合的效果,而在高濃度的溶壁酶區(qū)域,0.5%崩潰酶的輔助效果優(yōu)于1.0%崩潰酶的輔助效果。在復(fù)合酶液中,0.5%崩潰酶和2.0%溶壁酶的組合最好,原生質(zhì)體得率最高可達(dá)1.49×107個(gè)/mL。
2.1.2 酶解溫度對(duì)桑黃原生質(zhì)體制備的影響
溫度不同,酶活性不同,溫度是影響酶活的重要因素。溫度也會(huì)影響菌絲體的生理狀況。不同真菌的最適酶解溫度不同。在上述選定的酶系統(tǒng)和酶濃度的條件下,研究不同的酶解溫度對(duì)桑黃原生質(zhì)體制備的影響, 結(jié)果如圖4 所示。
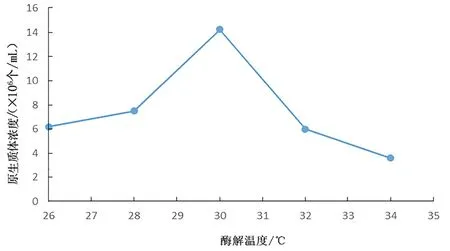
圖 4 酶解溫度對(duì)桑黃原生質(zhì)體制備的影響Fig.4 Effects of enzymolysis temperature on protoplast preparation of Phellinus igniarius
從圖4 中可以看出,酶解溫度對(duì)桑黃原生質(zhì)體的制備有較大影響。在一定程度上,酶解溫度的升高導(dǎo)致桑黃原生質(zhì)體得率不斷增加,當(dāng)酶解溫度為30℃時(shí),桑黃原生質(zhì)體得率為1.43×107個(gè)/mL,達(dá)到最高;當(dāng)酶解溫度超過(guò)30℃,桑黃原生質(zhì)體得率不斷降低。
2.1.3 酶解時(shí)間對(duì)桑黃原生質(zhì)體制備的影響
在上述選定的酶系統(tǒng)、酶濃度和酶解溫度的條件下,研究不同的酶解時(shí)間對(duì)桑黃原生質(zhì)體制備的影響,見(jiàn)圖5。
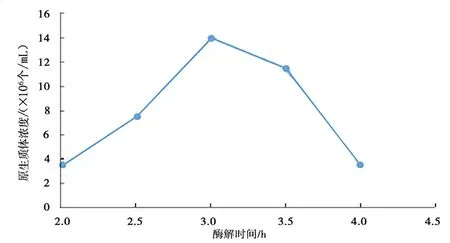
圖5 酶解時(shí)間對(duì)桑黃原生質(zhì)體制備的影響Fig.5 Effects of enzymolysis time on protoplast preparation ofPhellinus igniarius
由圖5 可知,原生質(zhì)體得率隨酶解時(shí)間的增加先增大后降低。當(dāng)酶解3.0 h 時(shí),原生質(zhì)體濃度最高。分析原因可能是酶解時(shí)間過(guò)短(小于3.0 h),桑黃菌絲酶解不夠充分;酶解時(shí)間過(guò)長(zhǎng)(大于3.0 h),則可導(dǎo)致原生質(zhì)體因酶解過(guò)度而破裂,原生質(zhì)體被破壞的速率大于原生質(zhì)體釋放的速率,故原生質(zhì)體濃度下降。
2.2 響應(yīng)面優(yōu)化試驗(yàn)結(jié)果
對(duì)表1 中的數(shù)據(jù),采用了 Design Expert 8.0.6 軟件進(jìn)行多元回歸擬合,以期進(jìn)一步考察各因素及其交互作用對(duì)桑黃原生質(zhì)體制備的影響,得出桑黃原生質(zhì)體濃度(Y)對(duì)酶解溫度(A)、酶解時(shí)間(B)、酶系統(tǒng)中溶壁酶濃度(C)的二次多項(xiàng)回歸模型方程為:
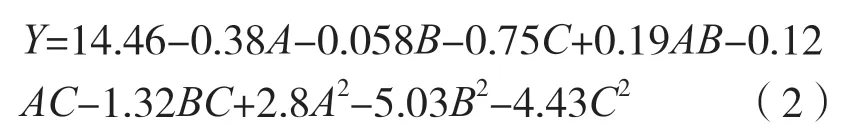
回歸方程顯著性檢驗(yàn)見(jiàn)表2。
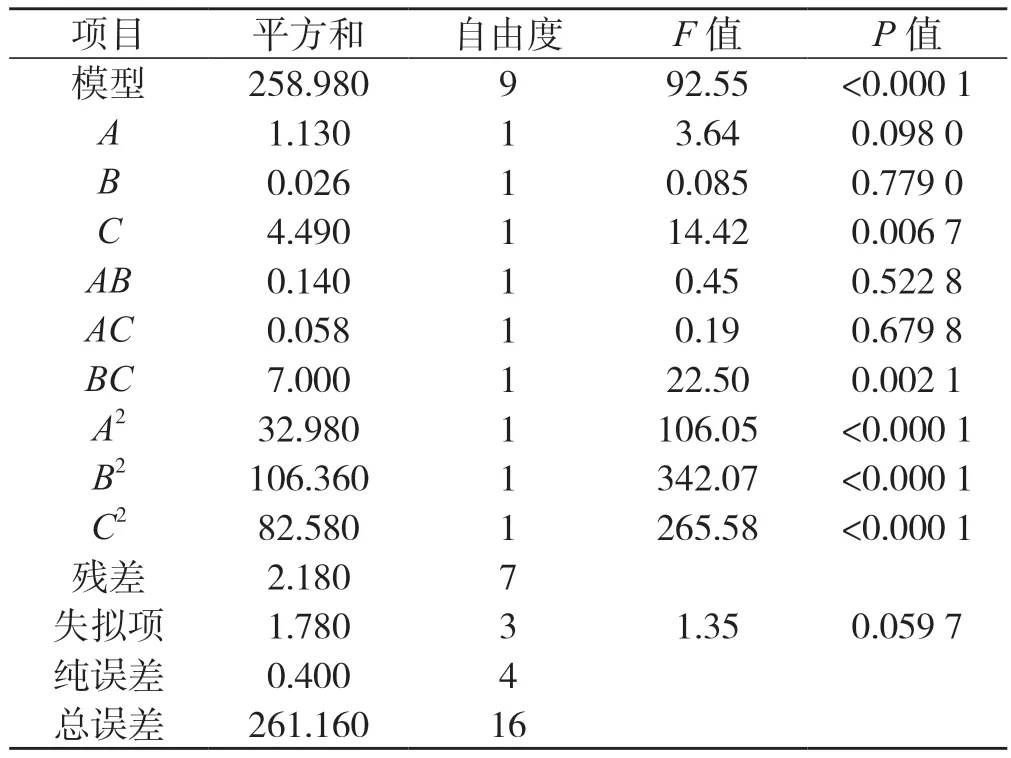
表2 回歸方程顯著性檢驗(yàn)Tab.2 Test of the significance of the regression equation
從表2 可以看出,模型的F=92.55、P<0.000 1,表明模型對(duì)桑黃原生質(zhì)體制備的影響顯著。R2=0.991 7,RSD=6.44,R2adj=0.981 0,可知模擬程度和模型相關(guān)度良好,且誤差較小。通常采用F檢驗(yàn)來(lái)判定各變量對(duì)響應(yīng)值影響的顯著性,如P值越小,則表示變量的顯著性越高。由表2 可知,方程中C、BC、A2、B2、C2影響較為顯著,結(jié)果表明試驗(yàn)因子的交互作用對(duì)響應(yīng)值的影響相對(duì)較小,不是簡(jiǎn)單的線性關(guān)系,二次項(xiàng)對(duì)響應(yīng)值也有較大的影響,這和模型回歸中的平方項(xiàng)影響顯著相對(duì)應(yīng)。
采用 RSM 研究各因子對(duì)桑黃原生質(zhì)體制備的影響,并結(jié)合 Box-Behnken 做響應(yīng)面圖,以考察溶壁酶的濃度、酶解溫度與酶解時(shí)間的交互作用對(duì)桑黃原生質(zhì)體制備的影響,進(jìn)而優(yōu)化桑黃原生質(zhì)體的制備條件。不同因素間交互作用的 3D 響應(yīng)曲面見(jiàn)圖 6~圖8 所示。
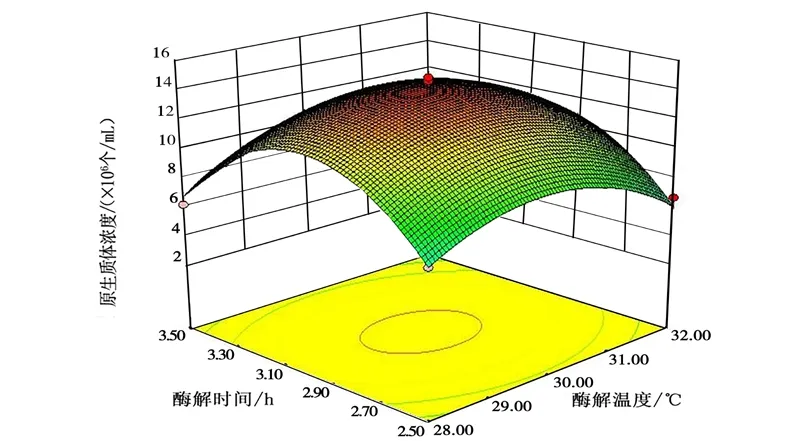
圖6 酶解溫度與酶解時(shí)間的交互作用對(duì)桑黃原生質(zhì)體制備的影響Fig.6 Effects of interaction between enzymolysis temperature and enzymolysis time on protoplast preparation of Phellinus igniarius
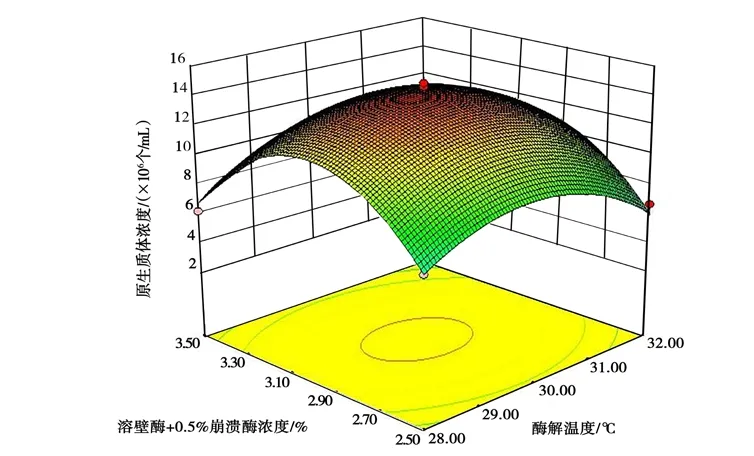
圖 7 溶壁酶濃度與酶解溫度的交互作用對(duì)桑黃原生質(zhì)體制備的影響Fig.7 Effects of interaction between lywallzyme concentration and enzymolysis temperature on protoplast preparation ofPhellinus igniarius
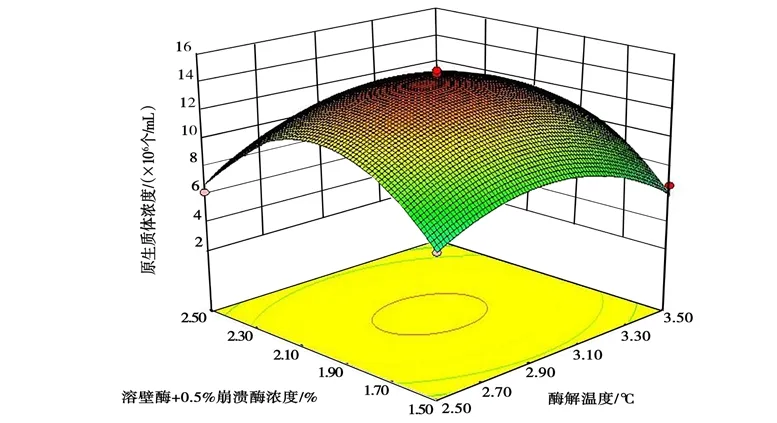
圖8 溶壁酶濃度與酶解時(shí)間的交互作用對(duì)桑黃原生質(zhì)體制備的影響Fig.8 Effects of interaction between lywallzyme concentration and enzymolysis time on protoplast preparation of Phellinus igniarius
從圖6~圖8 中可以清楚地看出,有關(guān)各因子之間交互作用的3D 圖。結(jié)果表明,試驗(yàn)范圍內(nèi)存在極值,不僅作為響應(yīng)面的最高點(diǎn),也是等值線最小橢圓的中心點(diǎn)。其中B、C的交互作用較強(qiáng),當(dāng)B和C的編碼值增大,響應(yīng)值(Y)卻呈減小的趨勢(shì)(圖8),分析原因可能是溶壁酶濃度過(guò)大、酶解時(shí)間過(guò)長(zhǎng),對(duì)原生質(zhì)體造成傷害;溶壁酶濃度過(guò)小、酶解時(shí)間過(guò)短,不能充分酶解菌絲得到原生質(zhì)體。從圖6 中可以看出,A與B的交互作用不強(qiáng),B(編碼值為0 以上)在A一定范圍內(nèi)均可獲得較大的響應(yīng)值,這與ANOVA 分析的結(jié)果吻合。圖7 中表明,A、C的交互作用也表現(xiàn)出和圖6 類(lèi)似的現(xiàn)象。由SAS 分析得到最大響應(yīng)值(Y) 時(shí)A、B、C對(duì)應(yīng)的編碼值分別為29.87℃、3.0 h、2.0%。與其相對(duì)應(yīng)的桑黃原生質(zhì)體制備的最佳條件為酶解溫度29.87℃,酶解時(shí)間3.0 h,溶壁酶濃度1.96%,理論最佳原生質(zhì)體得率為1.45×107個(gè)/mL。為檢驗(yàn)響應(yīng)面法的可行性,以最佳制備條件進(jìn)行桑黃原生質(zhì)體制備的驗(yàn)證試驗(yàn),考慮到操作的便利性,以酶解溫度30℃、酶解時(shí)間3.0 h、溶壁酶濃度2.0%為最佳。最終根據(jù)3 次平行試驗(yàn),獲得了實(shí)際的桑黃原生質(zhì)體平均得率為1.43×107個(gè)/mL,與理論值相差1.38%。因此,響應(yīng)面法對(duì)桑黃原生質(zhì)體制備的優(yōu)化是可行的,得到的桑黃原生質(zhì)體制備條件具有實(shí)際應(yīng)用價(jià)值。
2.3 桑黃原生質(zhì)體的再生
本試驗(yàn)研究了硫酸鎂、甘露醇、氯化鈉等3 種滲透壓穩(wěn)定劑對(duì)桑黃原生質(zhì)體再生的影響,按照1.5中的計(jì)算方法統(tǒng)計(jì)再生結(jié)果,再生率分別為0.67%、0.17%和0,桑黃原生質(zhì)體的再生菌落情況,見(jiàn)圖9。
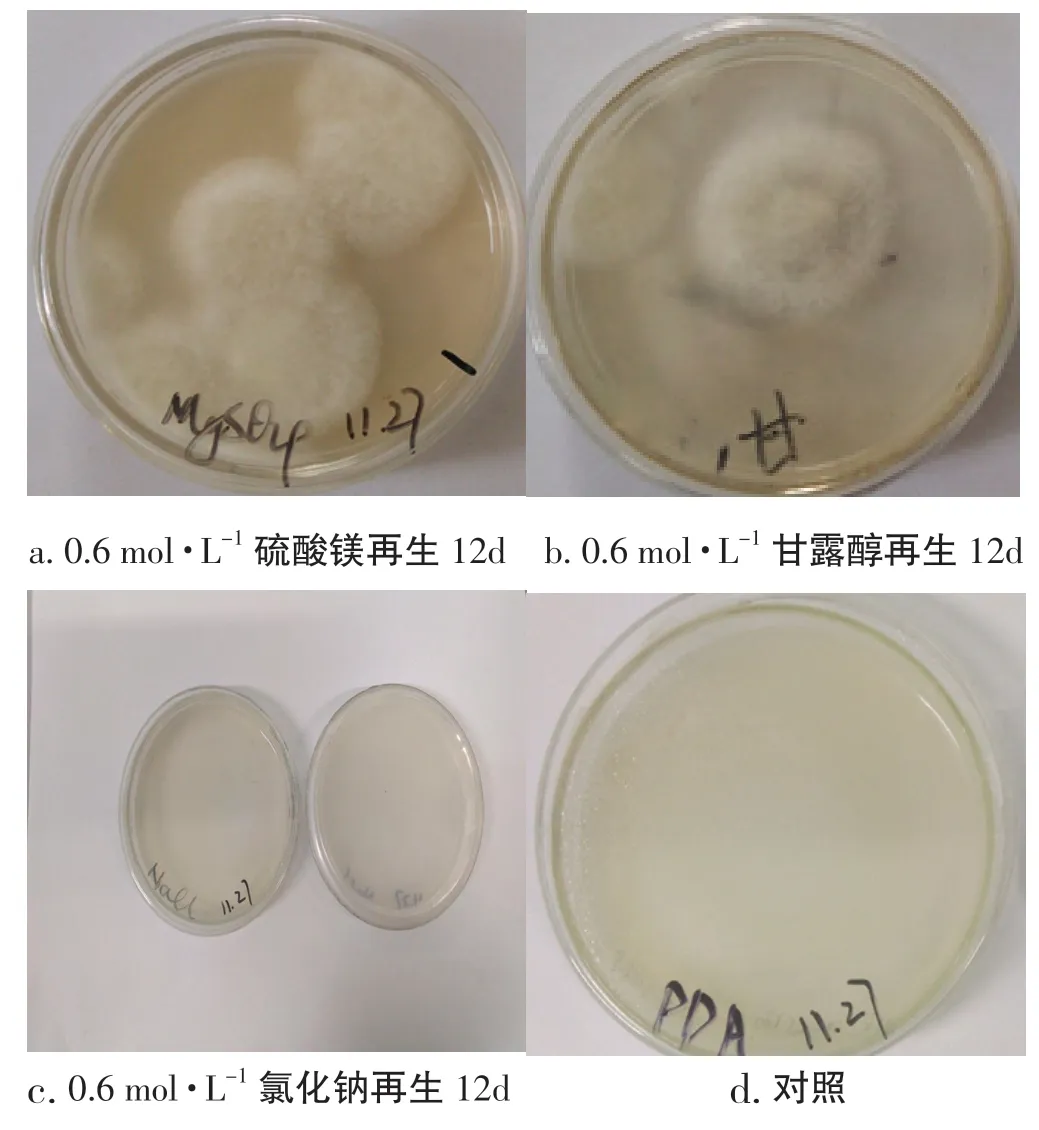
圖9 不同滲透壓穩(wěn)定劑對(duì)桑黃原生質(zhì)體再生的影響Fig.9 Effects of different osmotic stabilizer on protoplast regeneration of Phellinus igniarius
從圖9 試驗(yàn)結(jié)果可以看出,在3 種再生培養(yǎng)基中,添加了硫酸鎂的PDA 再生培養(yǎng)基較適于桑黃原生質(zhì)體的再生,在平板上長(zhǎng)出的菌落較多,且長(zhǎng)勢(shì)較強(qiáng);最差的是添加了氯化鈉的PDA 再生培養(yǎng)基,鮮見(jiàn)再生菌落,由此可知,在上述3 種滲透壓穩(wěn)定劑中添加了硫酸鎂的PDA 再生培養(yǎng)基較適合桑黃原生質(zhì)體的再生。
3 討論
隨著桑黃的各種藥理活性功能不斷被報(bào)道,人們對(duì)桑黃的需求與日俱增,然而受自身生理生化及環(huán)境條件的制約,野生桑黃資源稀缺,且有效藥用成分的含量較低,這迫使人們需要通過(guò)現(xiàn)代育種技術(shù)選育出高產(chǎn)活性成分的桑黃菌株。
原生質(zhì)體育種是大型真菌改良育種中一種很重要的方法,目前應(yīng)用較廣。因此,獲得高質(zhì)量的原生質(zhì)體對(duì)后期遺傳轉(zhuǎn)化尤為重要。自1972 年,裂褶菌原生質(zhì)體的成功分離后,大型真菌原生質(zhì)體的制備研究拉開(kāi)序幕[16]。相關(guān)學(xué)者的研究表明,原生質(zhì)體得率的影響因素主要包括酶解溫度、酶解時(shí)間、酶種類(lèi)、滲透壓穩(wěn)定劑等[17],其中使用復(fù)合酶的效果要好于單一酶[18-22]。
本研究在桑黃原生質(zhì)體的制備過(guò)程中,對(duì)酶及酶解條件等因素進(jìn)行了詳細(xì)研究,結(jié)果表明,使用溶壁酶與崩潰酶的混合酶系進(jìn)行酶解的原生質(zhì)體得率高于使用單一的溶壁酶。單因素試驗(yàn)結(jié)果展示了各個(gè)因素的單獨(dú)作用對(duì)桑黃原生質(zhì)體制備的影響。低濃度的酶液得到較小的原生質(zhì)體得率,隨著濃度的升高,原生質(zhì)體得率增加,但過(guò)高濃度的酶液會(huì)破壞原生質(zhì)體,使得原生質(zhì)體被破壞的速率大于原生質(zhì)體釋放的速率,故酶液濃度高時(shí),原生質(zhì)體得率下降。研究酶解溫度、酶解時(shí)間時(shí)也發(fā)現(xiàn)同樣的現(xiàn)象。此外,從響應(yīng)面試驗(yàn)結(jié)果分析中可以看出溶壁酶濃度、酶解溫度、酶解時(shí)間3 個(gè)因素對(duì)桑黃原生質(zhì)體制備的影響大小,其中溶壁酶濃度的影響最為顯著,其次是酶解溫度,最后是酶解時(shí)間。
原生質(zhì)體的制備與再生是2 個(gè)相互聯(lián)系且獨(dú)立的過(guò)程。在桑黃原生質(zhì)體的制備中,使用了甘露醇作為滲透壓穩(wěn)定劑,而在再生試驗(yàn)中篩選到的最適滲透壓穩(wěn)定劑是硫酸鎂,故再生滲透壓穩(wěn)定劑不一定是制備原生質(zhì)體時(shí)的滲透壓穩(wěn)定劑。試驗(yàn)中還發(fā)現(xiàn)添加了硫酸鎂的PDA 再生培養(yǎng)基在硫酸鎂濃度較高(0.6 mol·L-1)時(shí)瓊脂不易充分溶解,靜置后有大量沉淀,造成冷卻后培養(yǎng)基不會(huì)凝固或者凝固后仍有較大流動(dòng)性的現(xiàn)象,嚴(yán)重妨礙試驗(yàn)的進(jìn)行,這點(diǎn)和李剛等[20]的研究一致,但在硫酸鎂濃度較低(0.3 mol·L-1)時(shí)瓊脂可以充分溶解,溶液表現(xiàn)均勻一致,冷卻后的培養(yǎng)基也可以很好地凝固。我們?cè)诎l(fā)現(xiàn)瓊脂的這一特性后嘗試先在1 L 的培養(yǎng)基中溶解瓊脂,再加入0.6 mol·L-1硫酸鎂,結(jié)果發(fā)現(xiàn)培養(yǎng)基均勻一致,也可以很好地凝固,解決了李剛等人在使用硫酸鎂作為再生滲透壓穩(wěn)定劑時(shí)所遇到的培養(yǎng)基不能凝固的問(wèn)題。
最后,本研究發(fā)現(xiàn)桑黃原生質(zhì)體的再生周期較長(zhǎng),在7 d 以后才能肉眼觀察到再生菌落,原因可能是再生培養(yǎng)基的營(yíng)養(yǎng)不夠充分。對(duì)于如何加強(qiáng)再生培養(yǎng)基的營(yíng)養(yǎng),使桑黃原生質(zhì)體的再生更加迅速,長(zhǎng)出的的菌絲更加健壯,還需要進(jìn)一步詳細(xì)研究。
