藏母羊妊娠期日糧蛋白水平對初生羔羊GH 基因表達的影響
2019-12-19 08:58:06姜洋洋侯生珍王志有
飼料工業 2019年23期
姜洋洋 侯生珍 王志有
(青海大學農牧學院,青海西寧810000)
GH基因是提高動物的生長發育和生理活動的調節基因,不同的動物或同一種動物在不同生產水平下生長速度有顯著差異,這與其生長激素基因的表達水平緊密相關。GH基因生物學功能是促生長,動物通過自己的生長軸內分泌系統,從下丘腦直接分泌生長激素,釋放激素和生長抑素到垂體,促進生長激素的分泌,通過血液循環和肝臟表面的生長激素受體結合啟動細胞內信號傳導機制,通過對二聚體的構象變化,促進肝臟器官的胰島素生長[1-2]。生長激素是整個生長軸上最重要的激素,是動物生長的關鍵調控因子,刺激骨骼和軟骨的發育,加速脂肪分解代謝,促進動物的生長發育[3-4]。生長激素調節三大營養物質代謝,糖的轉化和氧化,蛋白質合成和氨基酸運輸,脂肪組織的合成需要酶的協助,生長激素可能會干擾脂肪酶的合成,轉錄抑制酶的活性,從而降低脂肪組織的合成速度[5]。從分子營養的角度來看,動物個體對營養物質的需求是不同的,動物對營養物質的消化、吸收和利用是否受到基因表達和調控的影響,研究不同蛋白質水平對綿羊生長發育及GH分泌、基因表達的作用,闡明飼料中蛋白質水平對綿羊生長發育及GH分泌、基因表達的影響具有重要意義。
1 材料與方法
1.1 試驗時間與地點
試驗時間:2018年8月~2019年3月。
試驗地點:試驗母羊妊娠前期放牧飼養地點在剛察縣沙流河鎮尕曲村,妊娠后期舍飼飼養地點在海東市樂都金元牧業有限公司。
1.2 試驗動物及試驗設計
試驗選擇60 只年齡、胎次、體況相近的藏母羊;對試驗母羊同期發情處理,確保在同一時間發情期受胎;母羊妊娠前期(0~85 d)在天然草場放牧飼養;妊娠后期(86~150 d)將母羊隨機分為2 組,每組30 只,進行全舍飼飼養,分別飼喂消化能相同而蛋白水平為7.5%和13.5%的日糧。母羊分娩后,每組選取3 只初生羔羊進行屠宰,采集瘤胃、網胃、瓣胃、皺胃、十二指腸、空腸、回腸、心臟、肝臟、肺臟、腎臟、肌肉等組織樣,樣品液氮保存待測。
1.3 飼養管理
試驗母羊采用前列腺醇同期發情處理(1 ml/只,2次,間隔12 d),第二次處理后按1∶10的比例投放種公羊進行集中配種。母羊妊娠前期(0~85 d)然草場放牧飼養,放牧草場為高寒草甸草場,早晨7:00 出牧,晚上20:00歸牧。母羊妊娠后期(86~150 d)全舍飼飼養,試驗開始之前對圈舍進行消毒處理,試驗母羊于每天早上8:00和下午18:00飼喂全混合日糧2次,自由采食,自由飲水;每天對料槽和水槽進行清洗,圈舍每周進行消毒,保持圈舍衛生、干燥、通風;試驗羊定期進行接種防疫。
1.4 試驗飼糧
參照NRC(2013)綿羊營養需要量標準和我國《肉羊飼養標準》,結合藏羊體重和營養需要特點,進行藏母羊妊娠后期飼糧配方設計,飼糧組成及營養水平見表1。
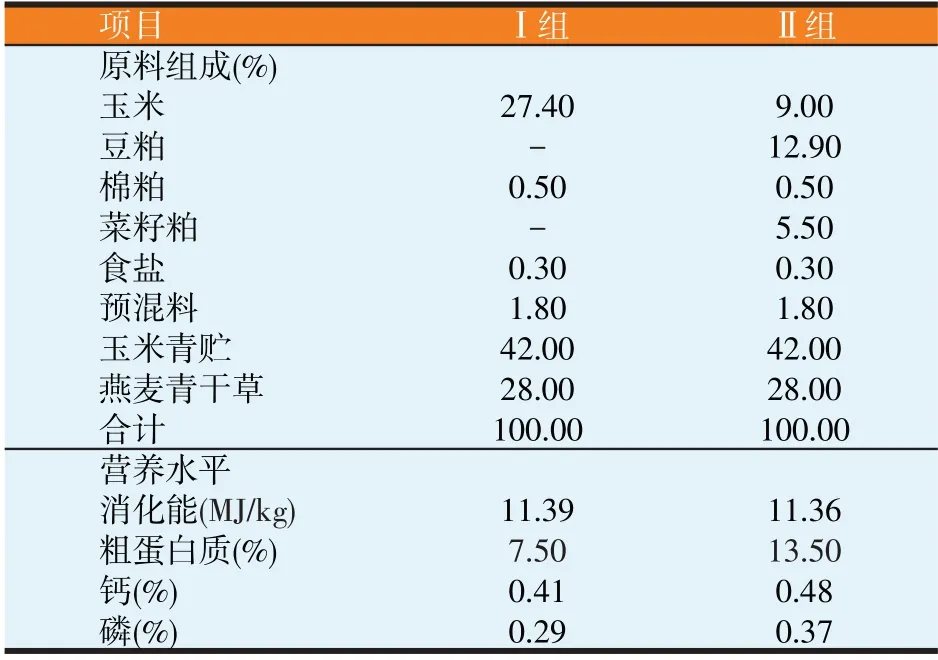
表1 飼糧組成及營養水平(干物質基礎)
1.5 測定指標及方法
1.5.1 總RNA的提取
將采集的組織樣品采用液氮研磨的方法充分粉碎,使用北京天根生化科技有限公司的總RNA 提取試劑盒并按照操作步驟提取樣品RNA。總RNA濃度和純度采用核酸定量儀進行檢測,確保總RNA 濃度在100 ng/μl 以上,A260/280 和A260/230 的OD 值在2~2.5 之間。1%的瓊脂糖凝膠電泳檢測RNA 的完整性或是否有蛋白質和DNA污染。
1.5.2 RNA反轉錄cDNA
利用北京天根生化科技有限公司FastKing RT kit cDNA的第一鏈合成試劑盒將提取的總RNA進行反轉錄。所有的操作都是在冰上進行的,實驗室設備必須先用RNase固相去除器進行處理。
1.5.3 引物的設計與合成
利用https://www.ncbi.nlm.nih.gov/gene/網站設計定量PCR 引物,確定引物序列,以GAPDH 為內參基因,由上海生工生物工程有限公司進行引物合成引物序列,產物長度及退火溫度如表2所示。
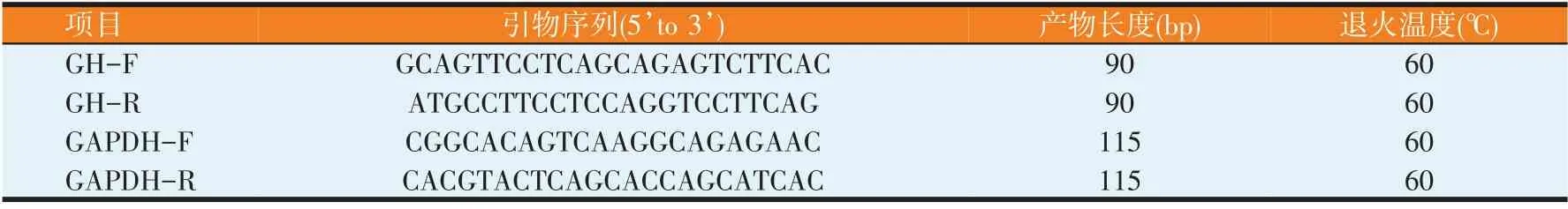
表2 引物信息
1.5.4 熒光定量PCR擴增
使用北京天根生化科技有限公司的SuperReal PreMix Color(SYBR Green)試劑盒,利用Rotor-Gene Q 實時定量PCR 儀測定目標基因在羔羊不同部位組織中的mRNA 表達量,每個樣品重復3 次,內參基因為GAPDH。反應條件中預變性、變性及退火的循環次數為40。具體反應體系和條件見表3和表4。

表3 PCR反應體系
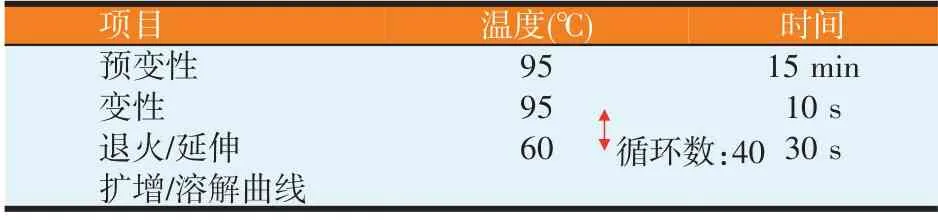
表4 PCR反應條件
1.6 統計分析
將試驗所得數據運用SPSS 20.0 軟件進行分析,結果采用“平均值±標準差”的形式表示。
2 結果
2.1 GH 基因在羔羊不同內臟器官及肌肉組織中表達量的比較
由表5和圖1可見,試驗Ⅱ組初生羔羊心臟、肝臟、肺組織中GH基因的表達量明顯高于Ⅰ組(P<0.05)。

表5 GH基因在羔羊不同內臟器官及肌肉組織中表達量
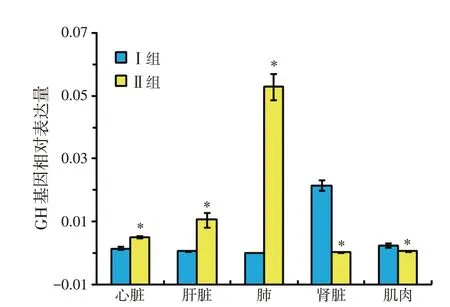
圖1 GH基因在羔羊不同內臟器官及肌肉組織中表達量
2.2 GH基因在初生羔羊不同胃腸道中表達量比較
由表6 和圖2 可見,初生羔羊瓣胃、皺胃、空腸中GH基因的表達量,試驗Ⅱ組顯著高于Ⅰ組(P<0.05),而瘤胃、十二指腸和回腸中GH基因的表達量,Ⅰ組和Ⅱ組間無顯著差異(P>0.05)。
3 討論
3.1 藏母羊妊娠后期日糧蛋白水平對初生羔羊內臟器官及肌肉GH基因表達量的影響
GH基因是調節生長和發育過程中生長激素生長和分泌的關鍵生長因子,具有促進肌肉和骨骼生長,促進動物生長發育,提高飼料報酬的作用。GH 基因存在于羔羊各個內臟器官和胃腸道及肌肉組織,其表達有顯著差異。章玲玲等[6]對家禽心臟、肝臟、脾臟、肺、腎臟和肌肉中GH 基因表達量進行了研究,證明GH 基因在肌肉組織表達量最低,肺中的表達量最高。張玲等[7]在研究不同日糧營養水平對番鴨GH基因在肌肉組織中表達情況的實驗中發現,在番鴨早期生長發育過程中,GH 基因表達會因日糧營養水平的不同而出現差異。本試驗結果表明,在兩種蛋白水平下,腎臟和肺中GH基因的表達量分別為最高和最低,這與前人的研究結果有相似之處,具有區別的原因可能是所選試驗物種的差異亦或營養水平設置梯度的差異。藏母羊妊娠后期日糧蛋白水平為13.5%時,初生羔羊心、肝、肺組織中GH 基因的表達量明顯高于7.5%蛋白組(P<0.05),說明妊娠后期提高藏母羊日糧蛋白水平能顯著提高胎兒內臟和肌肉組織的生長發育,此結果與張玲的研究結果一致。

表6 GH基因在羔羊不同胃腸道中表達量
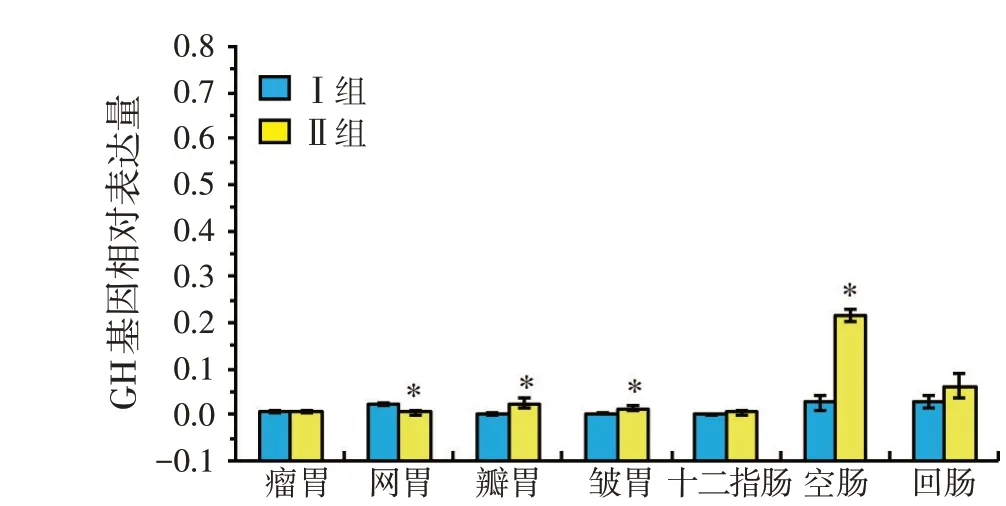
圖2 GH基因在羔羊不同胃腸道中表達量
3.2 藏母羊妊娠后期日糧蛋白水平對初生羔羊胃腸道GH基因表達量的影響
腸道的生長發育受多種因素的影響,如遺傳、養分、環境和神經內分泌等。腸道具有兩方面作用,既作為消化和吸收養分物質的重要器官,又是保持機體內環境穩定的先天性屏障。十二指腸是重要的消化器官,擔負著消化和吸收機體所需的大多部分養分物質的功能。因此,十二指腸發育的狀況在很大程度上決定動物的生長速度[8]。殷新平[9]對黃顙魚生長激素基因全長進行克隆測序,分析GH 在組織中的差異性表達,結果表明GH基因只在垂體中特異表達,而在肌肉、脾臟、脂肪、腎臟、胃、盲腸、心臟、頭腎、精巢、腸、鰓、肝臟、魚皮和腦這14 個組織中均未檢測到表達。曹運長等研究表明,垂體中GH 表達量與腎臟相比差異極顯著,垂體GH 基因表達量較高與藍太陽魚表達結果一致,在胃中檢測不到表達。胃的生長發育也離不開GH的調節,前人實驗已相繼表明在大鼠、兔子和人的胃中有GHr 基因的表達[10]。Nagano M(1995)用RT-PCR 結合Southern 雜交的方法也檢測出在人、大鼠和兔子胃組織中有GHr 的表達,以上研究結果表明,GH 對胃的生長有重要的調節作用。本試驗結果表明,藏母羊妊娠后期日糧蛋白水平為13.5%時,初生羔羊的瓣胃、皺胃、空腸GH基因表達量均顯著高于7.5%蛋白組(P<0.05),而瘤胃、十二指腸和回腸中GH基因的表達量,7.5%蛋白組和13.5%蛋白組間無顯著差異(P>0.05)。
4 結論
本試驗條件下,藏母羊妊娠后期日糧蛋白水平為13.5%時,初生羔羊心臟、肝臟、肺、瓣胃、皺胃、空腸中GH基因的表達量明顯高于7.5%蛋白組(P<0.05),妊娠后期提高藏母羊日糧蛋白水平能顯著提高胎兒內臟器官、肌肉組織和胃腸道的生長發育。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
火花(2019年12期)2019-12-26 01:00:28
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
人大建設(2019年12期)2019-05-21 02:55:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
