受體介導的1-磷酸鞘氨醇信號通路在局灶性腦缺血損傷中作用
2019-12-06 08:45:22李潔琦閔連秋
中風與神經疾病雜志 2019年11期
關鍵詞:小鼠
李潔琦,李 萌,閔連秋
1-磷酸鞘氨醇(Sphingosine 1 phosphate,S1P)在不同器官中生物學行為和功能主要由其5個特異性g蛋白偶聯受體(G Protein-Coupled Receptors,GPCRs)[1,2]介導產生。基于S1P受體開發的藥物芬戈莫德(FTY720)就是一種基于在磷酸化后非選擇性的與5個S1P受體亞型中的4個綁定后形成的一種藥物[3]。目前該藥物已經用于治療多發性硬化癥[4]。然而,到目前為止S1P的受體亞型在每種疾病類型中的具體作用機制尚不清楚。S1P1在腦缺血再灌注損傷中的致病作用表明,FTY720在該病中是通過作為S1P1功能拮抗劑的來實現治療作用的[5]。此外,有研究發現在腦缺血再灌注損傷發作后S1P3的mRNA水平升高[6],這說明S1P3可能是腦缺血再灌注損傷的另一致病因素,FTY720對腦缺血再灌注損傷的療效也有可能通過抑制S1P3來實現[7]。然而,目前還沒有明確S1P3在局灶性腦缺血再灌注損傷中是否參與了腦損傷過程,以及S1P3的致病或神經保護作用的報道[8]。因此,本研究擬通過構建大腦中動脈閉塞(middle cerebral artery occlusion,MCAO)小鼠模型,在小鼠腦缺血再灌注后給予S1P3拮抗劑CAY10444干預,評估各組動物腦損傷、腦梗死、神經功能缺損和神經細胞死亡等情況。
1 實驗材料和方法
1.1 實驗材料
1.1.1 實驗動物 雄性成年昆明小鼠(32 g±3 g,6 w)購置于錦州醫科大學實驗動物中心。在晝夜照明07:00-19:00(光)、溫度(22±2)℃、相對濕度(60±10)%環境條件下飼養適應環境1 w后,進行后續實驗。
1.1.2 主要試劑 CAY10444(賽默飛世爾中國);Brdu、rabbit anti-NF-κB(p65)、rabbit anti-Iba1、Donkey anti-BrdU、Mouse anti β-actin、DAB購于北京博奧深生物技術有限公司;神經元蛋白提取試劑及嵌合綠色熒光PCR試劑購于上海基康生物技術有限公司。
1.2 實驗方法
1.2.1 動物模型制備及分組 小鼠用異氟烷進行麻醉,3%濃度用來誘導,以1.5%濃度維持。通過腹側頸部切口分離右頸總動脈。從頸動脈分叉處向頸內動脈引入一根硅膠包裹的6-0手術線,結扎阻斷大腦中動脈。大腦中動脈閉塞30 min后,解開結扎的手術線,使大腦區域完全恢復血流處于再灌注狀態。在手術期間,采用保溫措施將動物的體溫維持在(37.0±0.5)℃。手術后,將小鼠置于鼠籠子里飼養,自由進食,按期更換墊料,最后處死動物取腦標本。實驗動物分為以下3組,每組10只:1.模型組(MCAO組):按照上述方法制備MCAO小鼠模型;2.藥物干預組(MCAO+CAY組):制備MCAO模型動物,動物腦缺血再灌注時將CAY10444分別以0.1 mg/kg、0.2 mg/kg和0.5 mg/kg的濃度給小鼠腹腔注射;3.假手術組(正常對照組):小鼠除不進行大腦中動脈結扎閉塞外,其他手術操作與模型組相同。
1.2.2 神經功能評分和腦梗死評估 采用改良神經功能缺損評分(modified neurological severity score,mNSS)量表評估功能神經受損程度,以確定MCAO后的運動、感覺、平衡和反射障礙等[9]。在神經功能評分后,采用斷頸法處死小鼠,迅速取出大腦,并在的小鼠大腦磨具刀組中切2 mm厚切片。大腦組織切片用含2%TTC的生理鹽水在37 ℃下孵化20 min。拍攝TTC染色的腦切片圖像,使用圖像分析軟件計算梗死體積。
1.2.3 組織學觀察與分析 MCAO術后1 d和3 d取腦組織標本進行組織學分析。小鼠采用異氟烷進行麻醉,通過心臟灌注冷的PBS和4%多聚甲醛混合液對腦組織進行固定。分離大腦組織,并在含30%蔗糖的灌流固定液中孵育,4 ℃冰箱過夜。將腦組織切成20 μm厚切片。在MCAO后1 d進行TTC組織化學染色。腦切片依次在梯度乙醇中水化(100%浸泡3 min,70%浸泡1 min,30%浸泡1 min),去離子水沖洗,0.06% KMnO4氧化。切片在0.1%醋酸溶液中,用0.01%的 TTC染色30 min,去離子水漂洗,在載玻片烘片機中干燥,二甲苯浸泡透明處理,然后蓋玻片。
1.2.4 Iba1免疫組織化學染色 在MCAO術后1 d和3 d行Iba1免疫組化染色。5 μm 厚切片,二甲苯脫蠟,酒精梯度入水,高壓鍋內枸櫞酸鹽進行抗原修復。切片在含1% H2O2的PBS中氧化15 min,含1%胎牛血清(FBS)的0.3% Triton-X100中阻斷1 h,阻斷非特異性蛋白結合。然后,腦切片與兔抗Iba1抗體(1∶500),在4℃ 孵育過夜。次日,切片與HRP標記的抗兔Ig-G二抗(1∶200)室溫下孵育1h。DAB顯色,鏡下觀察終止顯色。用下行乙醇梯度濃度脫水,二甲苯透明,封片。
1.2.5 BrdU / Iba1免疫熒光雙染 在MCAO后2 d和3 d,每隔12 h給予小鼠尾靜脈BrdU(50 mg/kg溶于PBS中)注射。小鼠處死后,取MCAO后大腦組織切片,與Tris溶液在100℃下孵育30 min進行抗原修復,Triton X-100中封閉1 h。然后與BrdU(1∶200)抗體在4℃孵育過夜。切片用HRP標記的二抗(1∶200)進行孵育。DAB顯色。染色的切片再次用PBS沖洗(3×5 min),再次進行阻斷,與抗體Iba1(1∶500)在4℃孵育過夜。然后用與Cy3(1∶1000)標記的二抗孵育,用抗熒光猝滅試劑進行封片。
1.2.6 免疫陽性細胞定量分析 從每個區域的不同部分拍攝3張照片,使用Adobe Photoshop CS3軟件進行圖像分析。并計算免疫陽性細胞的數量。然后,以單位面積(mm2)的免疫陽性細胞的平均數量進行表示。
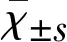
2 結 果
2.1 CAY10444對MCAO小鼠腦損傷和神經功能影響 在MCAO 24 h后,MCAO組小鼠在缺血的皮質和紋狀體區出現嚴重的腦梗死。使用S1P3拮抗劑CAY10444的干預組小鼠腦梗死體積明顯減少,且呈劑量依賴性(見表1)。給予最低劑量0.1 mg/kg的CAY10444干預組動物表現為無效,而給予0.2 mg/kg和0.5 mg/kg的CAY10444對減輕腦梗死有效,0.5 mg/kg組對減輕腦梗死和神經功能損傷療效最佳。通過mNSS分析發現,藥物干預組小鼠在運動、感覺、反射和平衡障礙等神經功能參數與MCAO組相比有顯著改善(見表1、圖1)。通過TTC染色分析缺血后24 h神經損傷程度,與MCAO組相比CAY10444藥物干預組減輕了神經損傷的程度(見表1、圖1)。
表1 各組動物腦梗死和mNSS評分表
注:MCAO 24 h后測定不同劑量CAY10444對梗死體積和神經功能的影響;與MCAO組比較**P<0.01,***P<0.001
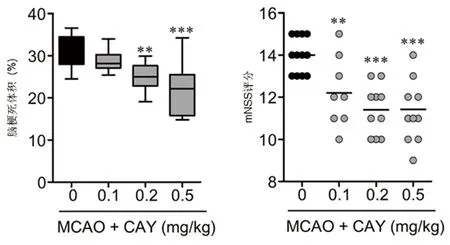
注:MCAO 24 h后不同劑量CAY10444對梗死體積和神經功能的影響。與MCAO組比較**P<0.01,***P<0.001
圖1 各組動物腦梗死組織學評估
2.2 CAY10444對MCAO腦內小膠質細胞活化和增殖的影響 MCAO后1 d和3 d通過Iba1免疫組化分析,發現MCAO組小膠質細胞的活化情況增強,表現為缺血半球的Iba1陽性細胞數量較假手術組增多。與MCAO組相比,CAY10444藥物干預組在上述兩個時間點上Iba1免疫陽性細胞的數量
顯著減少,且呈時間和區域依賴性,表現為藥物干預組小鼠大腦缺血周圍和缺血核心區域活化的小膠質細胞數量均明顯減少(見圖2、圖3)。此外,CAY10444藥物干預組小鼠大腦缺血核心區阿米巴樣形態的小膠質細胞數量顯著減少,即阿米巴形態小膠質細胞/分枝樣形態小膠質細胞比例降低(見圖3)。
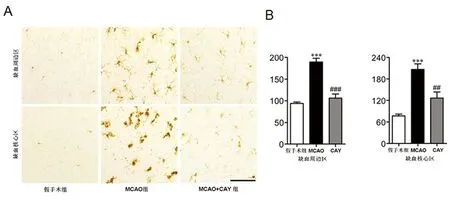
A:CAY干擾對MCAO后1 d小膠質細胞活化的Iba1免疫組織化學染色。P為缺血周圍區,C為缺血核區域。標尺=200 μm。B:缺血核心區和周邊區Iba1免疫陽性細胞數量的定量分析。與假手術組比***P<0.001。與MCAO組比##P<0.01,###P<0.001
圖2 腦缺血后1 d,CAY10444降低小膠質細胞活化情況
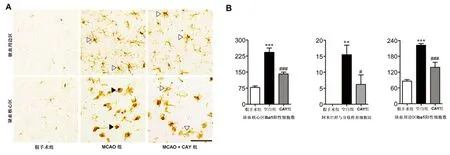
A:缺血周邊(P)和缺血核心(C)區域Iba1免疫陽性細胞圖。標尺=200 μm。B:缺血核心和周邊區Iba1免疫陽性細胞數量、Iba1陽性細胞的形態學改變(阿米巴樣/分支樣小膠質細胞)。與假手術組比較**P<0.01,***P<0.001;與MCAO組比較#P<0.05,###P<0.001
圖3 腦缺血后3 d小膠質細胞的活化情況
在MCAO 3 d 后對腦內BrdU和Iba1進行雙重免疫熒光染色,發現MCAO組大腦缺血區小膠質細胞增殖明顯,表現為BrdU/Iba1雙免疫陽性細胞較假手術組增多。與MCAO組相比,CAY10444藥物干預組BrdU/Iba1雙免疫陽性細胞的數量明顯減少(見圖4)。
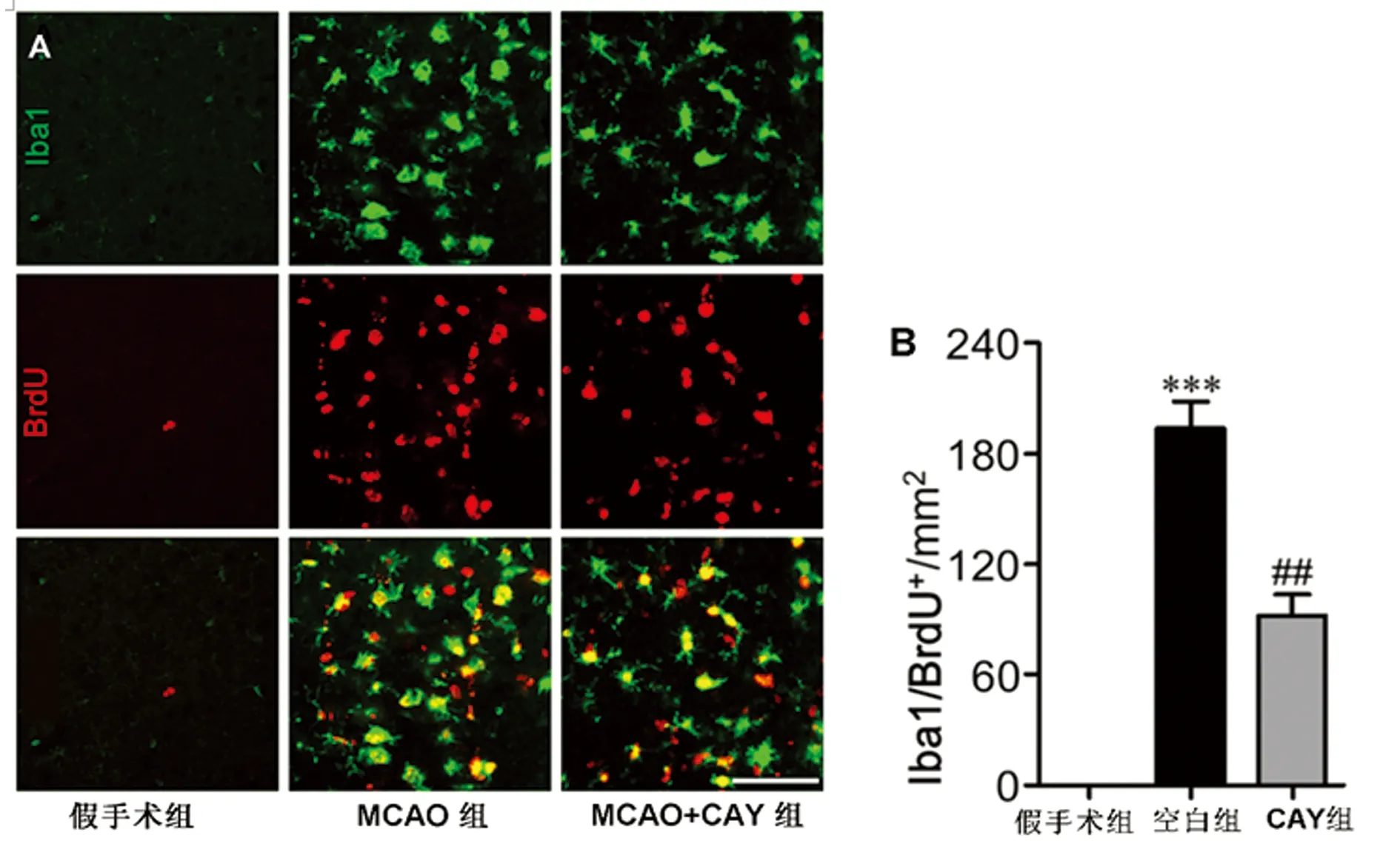
A:BrdU/Iba1雙免疫熒光法測定腦缺血后3 d,邊緣區BrdU/Iba1免疫雙陽性細胞圖。標尺=50 μm。B:BrdU/ Iba1免疫陽性細胞定量分析。與假手術組比較***P<0.001。與MCAO組比較##P<0.01
圖4 CAY10444對MCAO后3 d的小膠質細胞增殖的影響
3 討 論
在本研究中,我們探討了S1P受體的亞型S1P3在局灶性腦缺血中的致病機制,尤其是對小膠質細胞生物學的影響。MCAO后,通過特異性拮抗劑抑制S1P3活性可減輕腦損傷。S1P3在缺血性腦中的致病作用與小膠質細胞的活化密切相關,包括活化的小膠質細胞數量增多、形態轉化為阿米巴樣和小膠質細胞增殖。
人們通過FTY720的作用發現受體介導的S1P信號通路是腦缺血損傷的一種可行的治療方案,因為磷酸化后的FTY720是5個S1P受體亞型中的4個亞型的非選擇性調控子,在嚙齒動物中具有神經保護作用。盡管FTY720治療缺血性腦損傷取得了一定的成功,但究竟哪個或哪幾個S1P受體亞型是FTY720的實際介導的因子仍不確定。有研究者采用局灶性腦缺血大鼠模證明S1P1是局灶性腦缺血的致病因素[10]。本研究中,我們發現S1P3可能是介導腦缺血損傷的另一種S1P受體亞型。另外,最近有報道認為FTY720 的磷酸鹽可作為S1P1的功能拮抗劑發揮神經保護作用[11],這啟發我們猜測S1P3也可能對腦缺血損傷起作用。盡管到到目前沒有直接證據能證明這個猜測,但是一些研究發現了FTY 720 磷酸鹽可拮抗S1P3信號通路。考慮到FTY720磷酸鹽對S1P1和S1P3的抑制作用以及既往研究結果,因此,我們猜測FTY720對腦缺血損傷的療效可能是通過抑制S1P1和S1P3活性實現的。有研究證實S1P2不是FTY720-磷酸鹽的靶點,但是S1P2可通過破壞缺血腦中的血管完整性介導腦缺血損傷。因此,目前為止,人們已經確定三種S1P受體亞型為腦缺血的致病因素。然而,目前尚不清楚S1P受體亞型對腦缺血損傷的介導作用是否存在差異,是否還有其他S1P受體亞型如S1P4或S1P5參與了腦缺血損傷機制[12]。
S1P3在大腦中有明確的致病作用,但它在缺血條件下的作用是具有組織選擇性的。早期的關于S1P3在非神經缺血模型中的作用的研究報道存在著一些爭議。如在心臟中,S1P2和S1P3的缺失均可加重小鼠心肌梗死,支持S1P3的心臟保護作用[13]。然而,在腎臟中S1P3又與缺血后的組織損傷有關[14]。骨髓組織S1P3的缺失可減輕腎缺血/再灌注后的組織損傷,其表達缺失可降低促炎細胞因子的表達水平,增加抗炎細胞因子的表達水平[15]。S1P受體的這些不同作用在S1P1中也存在。S1P1受體激動劑或內皮細胞S1P1缺失可減少或加重缺血后的腎損傷[16]。然而,在大腦中,S1P1的下調減輕了腦缺血后的腦損傷[17]。無論受體介導的S1P信號在非神經組織中的作用如何,S1P1、S1P2和S1P3均參與了腦缺血后的腦損傷發病過程。
在本研究中,由于CAY10444是廣泛應用的S1P3特異性拮抗劑,因此我們使用CAY10444作為S1P3拮抗劑來研究其在腦缺血中的作用[18]。有報道證明S1P2是腦缺血再灌注損傷的致病因素之一,因此S1P2不太可能介導CAY10444在腦缺血中的神經保護作用[19]。最后,腦缺血中CAY10444的療效有可能僅通過S1P3介導。
S1P3在腦缺血后的神經損傷作用似乎與腦內小膠質細胞的激活有關,因為腦卒中等幾種中樞神經系統(CNS)疾病都會發生類似的致病作用[20]。此前,通過體外和體內的研究發現,受體介導的S1P信號通路參與小膠質細胞的激活[21]。最近發現S1P3介導的局灶性腦缺血腦損傷主要通過小膠質細胞激活介導[22]。在本研究中,我們發現S1P3也與小膠質細胞的激活有關:使用S1P3特異性拮抗劑抑制S1P3活性,腦中活化的小膠質細胞的數量以時間和區域依賴的方式減少。此外,抑制S1P3活性降低了小膠質細胞的增殖。腦缺血組織中S1P3除了造成活化小膠質細胞數量增加外,還與活化的小膠質細胞的形態學轉化密切相關。在缺血再灌注后3 d時,缺血核心區域活化的小膠質細胞大多呈阿米巴樣,并主要通過釋放多種促炎介質導致缺血腦神經元損傷。我們的研究表明,抑制S1P3可顯著降低活化小膠質細胞向阿米巴樣形態轉化。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
