209株不同來源空腸彎曲菌分子特征分析
2019-10-15 10:41:12王聞卿李彩云崔琪奇朱林英
中國人獸共患病學報 2019年9期
趙 冰,黃 紅,王聞卿,李彩云,崔琪奇,朱林英
空腸彎曲菌(Campylobacter.jejuni)是世界上引起感染性腹瀉最常見的病原體之一[1]。人群中各個年齡段均易感[2]。在歐美發達國家,感染性腹瀉中的彎曲菌占很高比例[3-6]。空腸彎曲菌某些菌株能引起一些嚴重并發癥,例如吉蘭巴雷綜合征[7-8]。
菌株的分子分型對解釋傳染病的流行至關重要。傳統表型分型方法如血清分型以及生物分型分辨率不足。目前主流的分子分型方法如脈沖場凝膠電泳(PFGE)在短期暴發監測中顯示了很強分辨率,但是在長期監測中,對于遺傳關系的分析能力不足。基于序列分析為基礎的多位點序列分型(MLST)能夠靈敏地發現因種內和種間頻繁的水平基因交換引起的遺傳多樣性,其方法簡單,結果能在國際實驗室間比對,共享性好。其技術原理是對細菌的數個管家基因序列進行測序,數據庫將管家基因序列進行識別和組裝,生成序列型(Sequence Type,ST),應用生物信息分析軟件分析序列型。Dingle等人建立了空腸彎曲菌的MLST方案[9],并建立了成熟共享數據庫,該技術應用于空腸彎曲菌株間比對、溯源分析、遺傳進化分析[10-11]。
1 材料與方法
1.1 菌株的來源
1.1.1 人源株 選取浦東新區11家醫療機構作為監測點,依據系統抽樣原則,采集急性感染性腹瀉病人糞便標本,2014年共采集1 846件,2015年共采集2 100件。采集的糞便拭子標本接種Karmali瓊脂,42℃微需氧培養48 h,可疑菌株進行氧化酶、過氧化氫酶、馬尿酸水解試驗進行篩選,符合者再用熒光PCR試劑盒鑒定,通過分離共獲得144株空腸彎曲菌,其中2014年分離到70株,2015年分離到74株。1.1.2 禽源株 利用浦東新區設置的禽流感監測點,2014-2015年共采集樣本1 345件,樣本主要類型有禽類泄殖腔、禽類糞便、禽舍籠具、養殖場地面、污水等,檢測方法同1.1.1,通過分離共獲得56株空腸彎曲菌。
1.1.3 食品株 選取腹瀉病原譜監測點附近的超市、農貿市場,采集禽畜肉、豬內臟共252件。采集的食品取25 g剪碎后,加入100 mL TSB增菌液,混勻后取上清液50 mL,10 000r/min離心,取沉淀接種karmali瓊脂,以下過程同1.1.1,通過分離共獲得9株空腸彎曲菌。
1.2 儀器與試劑
1.2.1 主要儀器 PCR儀(AB9700)、Bio-Rad凝膠成像系統(GEL DPC)
1.2.2 主要試劑 PCR擴增試劑盒購自上海生工,DNA marker購自于大連寶生物,管家基因擴增引物由Invitrogen公司合成,10XTBE緩沖液購自于伯樂,瓊脂糖購自于上海生工,細菌基因組DNA提取試劑盒購自于北京天根生物科技公司,微需養產氣袋購自于Thermo Fisher,空腸彎曲菌雙色熒光pcr試劑盒購自于碩世生物科技有限公司。
1.3 管家基因位點選擇 根據彎曲MLST數據庫方案(http://pubmlst.org/campylobacter)確定空腸彎曲菌的7個管家基因:asp A、gln A、glt A、gly A、pgm、tkt、unc A。
1.4 管家基因擴增 挑取少許菌落,用商品化細菌基因組DNA試劑盒提取。反應程序為94℃變性30 s,50℃(unc A),55℃(asp A,glt A,gly A,pgm,tkt),60℃(gln A)退火30 s,72℃延伸60 s,共35個循環。擴增序列見表1。
1.5 測序 取PCR產物委托上海奕躍生物技術公司進行sanger一代雙向測序。測序序列見表1。
1.6 數據處理與分析 將測序拼接后管家基因序列提交至 MLST數據庫 (http://pubmlst.org/campylobacter),獲得7個管家基因號,整合后確定ST型和CC。應用BioNumerics 7.1進行比對分析。

表1 管家基因擴增與測序引物Tab.1 Primers for amplification and sequencing of housekeeping genes
2 結 果
2.1 MLST測序結果 所有209株空腸彎曲菌共獲得110種序列型,歸屬25種同源復合體。歸屬ST-21同源復合體菌株最多,共計31株,占比為14.8%(31/209),其中人源株占比9.7%(14/144),禽類養殖環境株占比30.4%(17/56);歸屬ST-353同源復合體共計28株,占比為13.4%(28/209),其中人源株占比16.7%(24/144),禽類養殖環境株占比7.1%(4/56);歸屬ST-464同源復合體共計25株,占比為11.9%(25/209),其中人源株占比10.4%(15/144),禽類養殖場環境株占比17.9%(10/56);無法歸屬同源復合體共42株,占比為20.1%(42/209),其中人源株占比20.1%(29/144),禽源株占比16.1(9/56),食品株占比44.4%(4/9)。人源株和養殖場環境株在主要流行種群種類上相似,僅僅在結構數量上有區別,見表2。
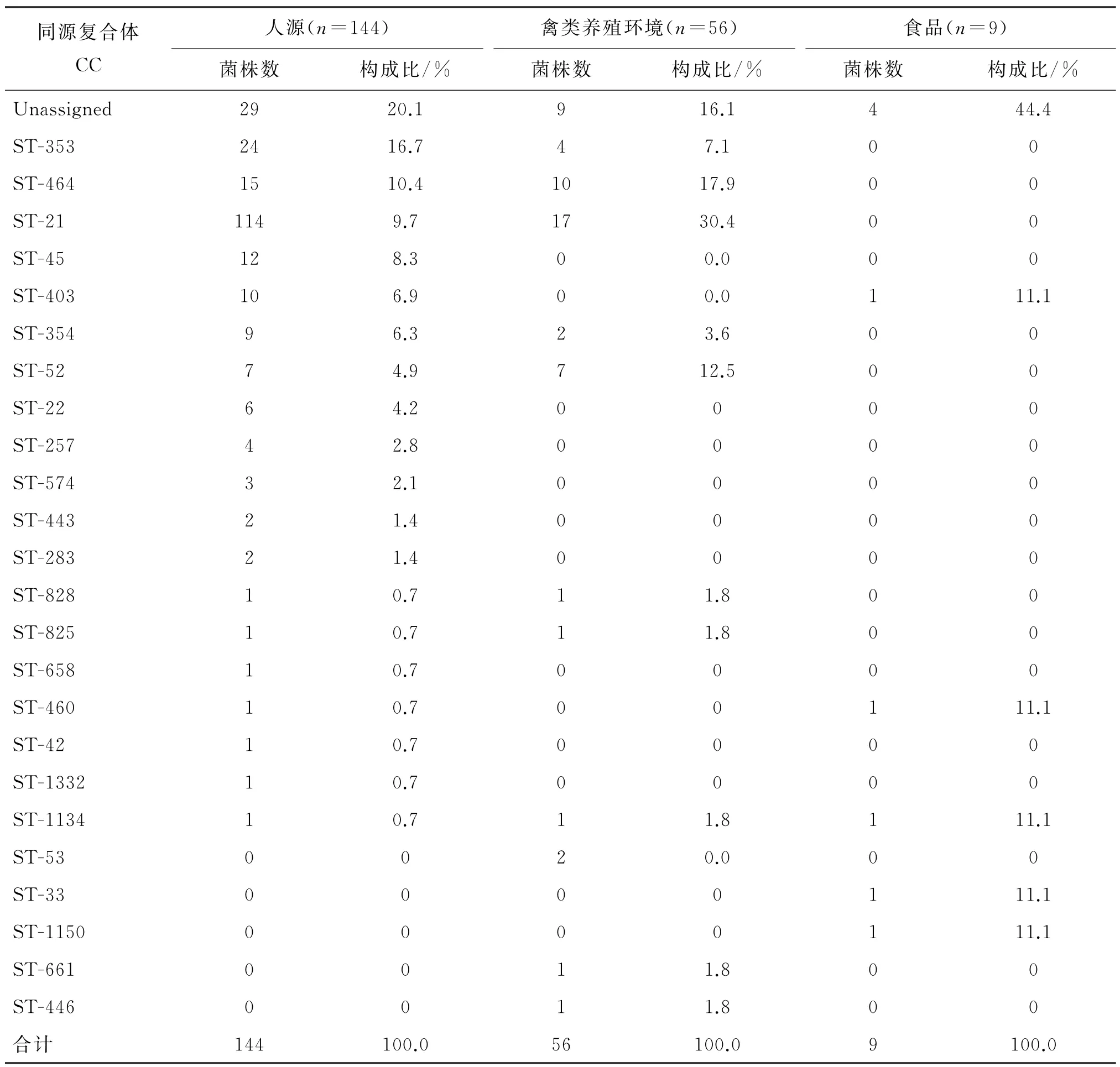
表2 不同來源菌株同源復合體所占比例Tab.2 The rate of isolates from different source belonging to various complex
2.2 MLST進化分析 用BioNumerics7.1繪制Neighbor Joining遺傳進化樹圖,見圖1,樹狀圖中清晰地展示菌株進化關系,圖中綠色代表2014年人源株,紅色代表2015年人源株,紫色代表禽類養殖環境株,黃色代表食品株。遺傳進化分析圖上我們總結成了5個主要聚類簇,聚類簇1是以ST45序列型為核心組成的ST-45CC,其規模較小,主要集中了來自腹瀉病人的菌株。聚類簇2是ST-50型為核心,ST-50隸屬于ST-21CC,是起源株ST-21型發展而來,監測數據中僅僅只有一株ST-21來自于腹瀉病人,ST50均為養殖環境株,人源株中未發現。圖1中央的包圍著ST-464形成了最大的一個聚類簇3,以ST-464起源命名的ST-464CC成為了最大的同源復合體。第4個聚類簇包含ST-354CC和ST-52CC,其規模較小,ST型多樣性,以人源株為主。第5個聚類簇是以ST-5為核心,其歸屬的ST-353CC該復合體也是全球和國內流行的序列型,在本次研究的樣本中,主要來自于腹瀉病人。食品株均呈散在分布,與禽源和人源株關系較遠。
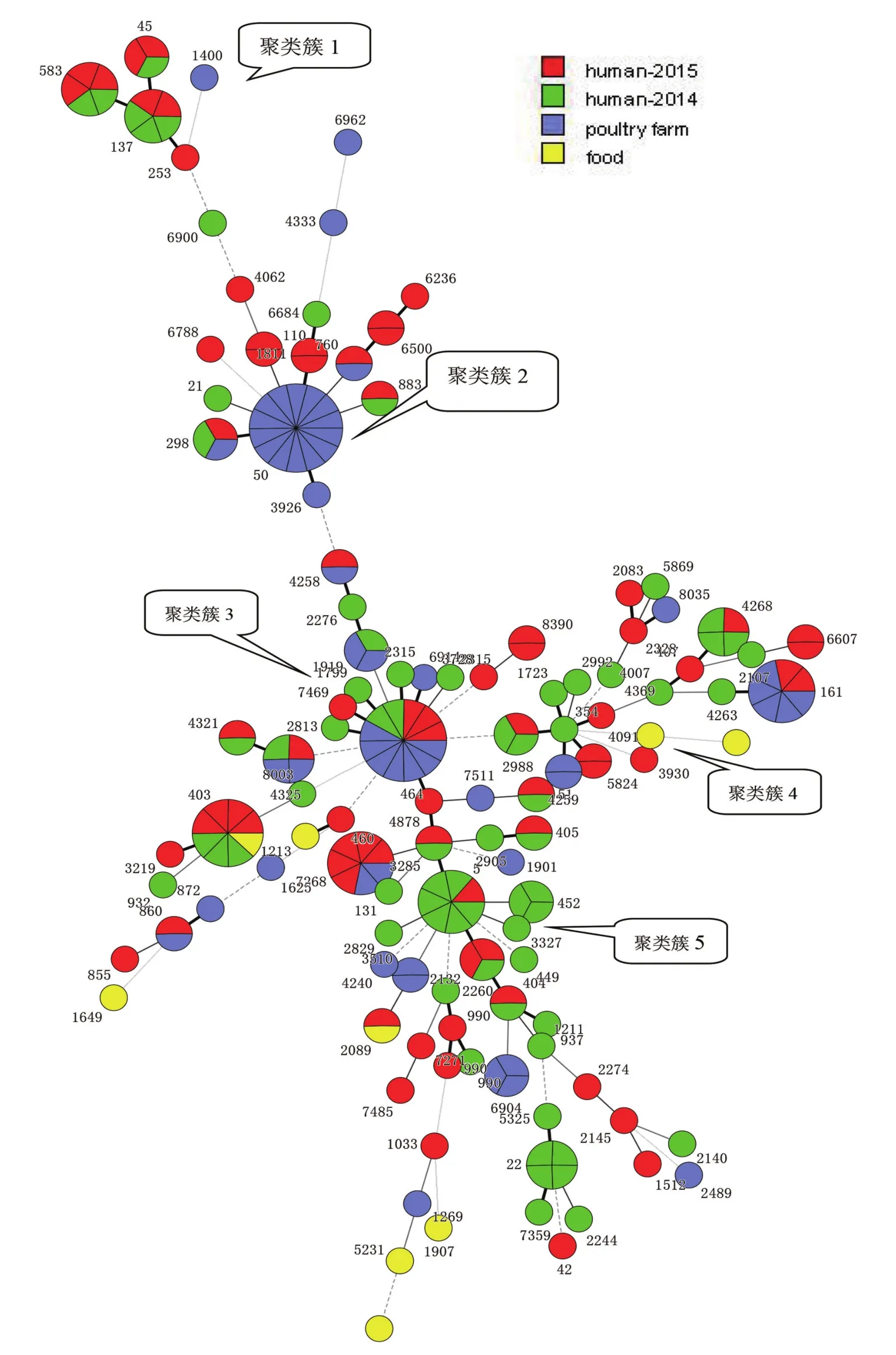
圖1 209株空腸彎曲菌遺傳進化樹Fig.1 Dendrogram of genetic relationships among 209 strains
3 討 論
空腸彎曲菌已經成為本地區主要的腹瀉病原體,但是過去一直缺乏簡便、分型能力強的方法,故無法對菌株進行更深層次的分析,MLST方法的應用,解決了難以分子分型的問題。MLST技術體現了高分辨率、分析能力強、共享性好的特點,在空腸彎曲菌分子分型中具有非常強的適用性。今后,可推廣成為多種致病菌來源、傳播途徑、進化分析、暴發識別的工具[9]。
本研究通過對209株不同來源的空腸彎曲菌進行MLST分析,得到了多種STs,雖然STs種類繁多,但是其中也具有較高的重復頻率,揭示了比較親密的遺傳關系。144株人源株分型結果顯示,本轄區人源株呈多樣性,從流行上分析,存在相對優勢的同源復合體,2014年的最優勢群體為ST-353CC(25.7%),2015年的最優勢同源復合體轉變為ST-21CC(14.6%),兩年的其他同源復合體分布呈多樣性,種類也較為相似,但結構發生變化。
從禽類養殖場所的環節陽性株分析結果看,ST-50檢出頻次最高的序列型,ST-50所屬的ST-21CC在華東地區禽類監測中普遍存在[12],而且在發達國家養殖業中也普遍流行[13-15],本轄區兩年的監測數據并沒有在腹瀉病人中發現ST-50序列型的蹤跡。從圖1進化關系中可以發現大多數所屬ST-21CC的人源ST型與ST-50有很近進化關系,有可能ST-50從禽類遷徙到人群以及人群間傳播過程中變異分化,本次研究由于數據有限,進化趨勢特征需要多年監測才能得到正確的判斷。ST-464是國內最為流行的序列型,本次研究共檢出12株,禽糞便和腹瀉病人中均能發現,其所屬的ST464CC,其家族更大,共有25株,占比為12.0%,在禽類糞便、養殖環節、腹瀉病人均有分布,這些數據揭示了本轄區人感染空腸彎曲菌與禽類關系非常密切。ST353CC也是全球和國內流行的克隆群[16],在本次研究的樣本中,主要來自于腹瀉病人,本研究沒有對外來的禽類進行大樣本量的分析,所以無法判斷是外源輸入還是本地流行。
本次研究食品株有44.4%(4/9)序列型無同源復合體歸屬,其主要原因本次研究的食品樣品數量和種類的相對缺乏,從食品中主要分離到的是與本研究無關結腸彎曲菌,共48株,而目標菌空腸彎曲菌僅僅9株,遺傳背景信息太少,無法反映食品株和人源株以及禽類養殖環境株的關系,也是本研究的一個遺憾之處。
本研究對菌株的序列型以及相互關系進行描述性分析,無法準確推測總體,僅能提出假設,若要推測群體間的確切的遺傳關系,下一步的重點工作需要進一步持續地對不同來源的菌株分子信息進行積累,應用分子信息模型工具量化分析不同群體間的遺傳關系遠近,并進行歸因分析,確切地鎖定最可能的危害因素,為控制該病原提供客觀的依據。
綜上所述,本地區的空腸彎曲菌感染以散發為主,序列型呈多樣性,種群結構相對穩定,禽源株的與人源株存在密切的關系,主要的流行群在本轄區相對穩定存在。研究結果提示我們,加強禽類養殖、運輸、加工的管理,持續對空腸彎曲菌的動態監測是控制和降低人群彎曲菌感染的重要途徑。
利益沖突:無
引用本文格式:趙冰,黃紅,王聞卿,等.209株不同來源空腸彎曲菌分子特征分析[J].中國人獸共患病學報,2019,35(9):865-869,874.DOI:10.3969/j.issn.1002-2694.2019.00.095
