乙酰輔酶A羧化酶在卵巢癌中的表達與促增殖作用研究
2019-08-24 07:42:12羅劍波鄧曉楊
轉化醫學雜志 2019年4期
鄧 潔,羅劍波,彭 聰,鄧曉楊
腫瘤是一種基因病,同時也是一種代謝性病[1-3]。大量研究表明,與正常細胞相比,腫瘤細胞代謝網絡發生了顯著的改變,主要體現為:糖酵解與脂肪酸從頭合成活性的顯著增強,以及谷氨酰胺代謝異常活躍[4]。靶向異常改變的細胞代謝網絡被認為是良好的腫瘤靶向治療策略[5-7]。目前,與糖酵解和谷氨酰胺代謝相比,腫瘤細胞的脂肪酸代謝異常受到的關注相對較少[8]。而事實上,腫瘤細胞的快速分裂增殖需要大量的脂肪酸為其生物膜等結構性成份的合成提供原料[9],脂肪酸代謝重編程已被證實在促進腫瘤細胞惡性增殖及轉移中發揮著關鍵的作用[10-11]。
乙酰輔酶A羧化酶(Acetyl-CoA carboxylas 1,ACC1)是參與脂肪酸合成調控第一步反應的關鍵酶,負責催化乙酰輔酶A羧化生成丙二酰輔酶A。近年來,ACC1被證實在多種類型的腫瘤細胞中表達異常升高,其中包括肝癌、肺癌、乳腺癌與胰腺癌等[12-15],以ACC1為靶點的抑制劑被證實可通過抑制脂肪酸合成而抑制腫瘤細胞的增殖[16-17]。然而,卵巢癌細胞中ACC1的表達與作用尚不十分清楚。本研究旨在探討ACC1在卵巢癌組織中的表達及其在卵巢癌細胞增殖中的調控作用。
1 材料與方法
1.1 卵巢癌細胞系與組織標本
1.1.1 卵巢癌細胞 卵巢癌細胞系SKOV3購自美國模式菌種收集中心細胞庫,細胞培養采用含10%胎牛血清(SV30087,Hyclone,美國)的RPMI-1640培養基(R8758,Sigma-Aldrich,美國),培養箱環境設為37 ℃含5%濃度的CO2。
1.1.2 臨床組織標本 共收集10例卵巢癌患者癌與癌旁組織,所有患者具有完整臨床資料與明確的病理診斷,并事先簽署了知情同意書,術中取得組織后立即置于液氮中保存。
1.2 實驗方法和步驟
1.2.1 實時熒光定量PCR(quantitative real-time PCR,qRT-PCR) 首先,用組織RNA提取試劑盒(R6688,美國OMEGA公司)對卵巢癌組織與癌旁組織中總RNA進行提取。隨后,用反轉錄試劑盒(RR037A,日本TAKARA公司)將所提RNA反轉錄合成cDNA。PCR實驗開始前需分別合成目的基因ACC1與內參基因GAPDH的引物,ACC1引物序列為:上游5′-ATGTCTGGCTTGCACCTAGTA-3′,下游5′-CCCCAAAGCGAGTAACAAATTCT-3′;GAPDH序列為:上游5′-ACAACTTTGGTATCGTGGAAGG-3,下游5′-GCCATCACGCCACAGTTTC-3′;用2-△△Ct相對定量法對最終結果進行計算。
1.2.2 Western Blot 首先,用含蛋白酶抑制劑的RIPA裂解液對卵巢癌癌組織與癌旁組織進行裂解以提取總蛋白,并采用BAC法對蛋白濃度進行檢測。隨后加入商品化的蛋白上樣緩沖液并在沸水中煮5 min,按BCA定量結果將相同量的蛋白上樣至聚丙烯酰胺凝膠孔中后即可進行電泳分離,待酚藍至凝膠底部時即可停止電泳。隨后將凝膠中蛋白用“三明治”法轉移至聚偏二氟乙烯膜表面。膜用5%的BSA溶液室溫封閉1 h后,加入稀釋過的ACC1抗體(1∶1 000)并于4 ℃冰箱中孵育過夜,PBS洗滌3次(每次5 min)后加入二抗并于28 ℃孵育2 h,繼續用PBS洗滌3次(每次5 min)。最后,用ECL發光系統對結果進行分析。
1.2.3 siRNA下調卵巢癌細胞SKOV3中ACC1表達首先由上海吉瑪公司合成兩條靶向ACC1不同區域的siRNA干涉片段,靶向序列分別為:siACC1#1為CCUACAAUGGGAACAGCUA;siACC1#2為GAACUUAACCGGAUGCGUA。隨后,將合成好的siRNA轉染SKOV3細胞,方法為:將對數生長期的SKOV3細胞以2×105個/孔的密度種至6孔板中,過夜培養后進行siRNA轉染,轉染步驟嚴格按脂質體(lipofectamine 2000,lip2000)操作說明進行。分別將lip2000與siRNA片段用不含血清的RPMI-1640培養液稀釋,靜止5 min后將二者混和并進行上下顛倒混合,靜置25 min后用移液器吸取100 μL并加入6孔板細胞中,隨后將細胞置于孵箱中培養,6 h后更換含血清培養基繼續培養24 h。此時,細胞即可分別用于四氮唑藍鹽[3-(4,5-diethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-etrazolium inner salt,MTS]增殖、5-乙炔基-2′-脫氧尿苷(5-ethynyl-2′-deoxyuridine,EDU)與凋亡實驗。
1.2.4 MTS增殖實驗 用“1.2.3”中所述的方法對卵巢癌細胞SKOV3中ACC1表達進行siRNA干涉處理,胰酶消化并離心收集細胞按3×103個/孔的密度接種至96孔板中。分別在第1、2、3、4、5天進行MTS檢測,具體操作步驟如下:用移液器將10 μL的MTS工作液加入96孔板中并在37 ℃反應4 h,隨后于450 nm處檢測吸光值(optical density,OD)。以時間(天)為橫軸,OD為縱軸繪制細胞的增殖曲線。
1.2.5 EDU實驗 用“1.2.3”中所述的方法對卵巢癌細胞SKOV3中ACC1表達進行siRNA干涉處理,胰酶消化并離心收集細胞按5×104個/孔的密度將細胞接種至共聚焦顯微鏡專用細胞培養皿中培養過夜后即可進行后續EDU染色實驗,染色過程嚴格按EDU試劑說明書(廣州銳博生物公司,貨號C10327-1)進行,細胞依次經Triton-X-100打孔、4%多聚甲醛固定后、EDU染色、細胞核復染后,用激光共聚焦顯微鏡對結果進行觀察。以時間(天)為橫軸,OD為縱軸繪制細胞的增殖曲線。
1.2.6 細胞凋亡的流式細胞術檢測 首先,用“1.2.3”中所述的方法對卵巢癌細胞SKOV3中ACC1表達進行siRNA干涉處理,胰酶消化并離心收集細胞,隨后按凋亡試劑盒說明(美國Everbright lnc公司,貨號Y6026)對細胞進行Annexin V與PI染色,最后上機進行細胞凋亡的流式細胞術檢測。
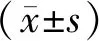
2 結果
2.1 卵巢癌患者癌與癌旁組織中ACC1的表達情況 分別用qRT-PCR與Western Blot對10例卵巢癌患者癌組織與癌旁組織中ACC1的mRNA與蛋白表達水平進行了檢測,結果如圖1A與1B所示:所有10例卵巢癌患者癌組織中ACC1表達均不同程度的高于癌旁組織(P<0.01),表明ACC1在卵巢癌細胞中的表達發生了顯著上調。
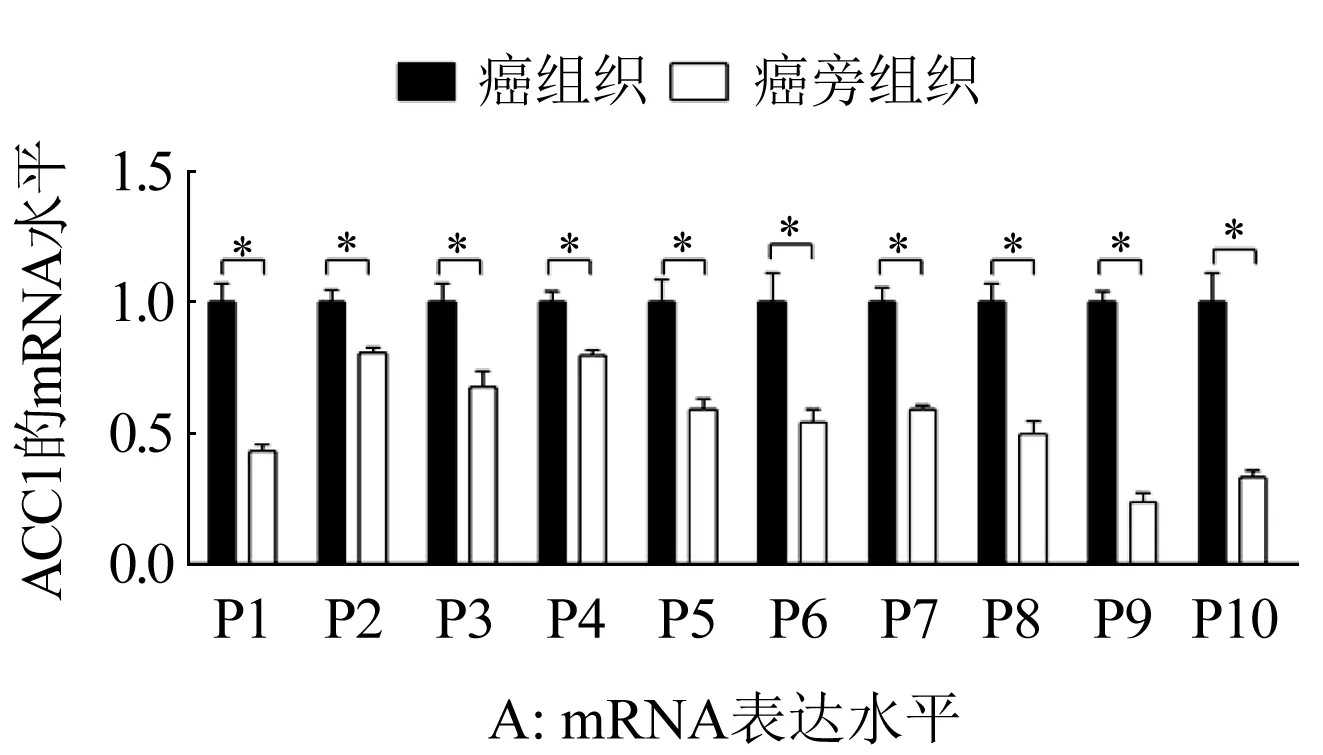
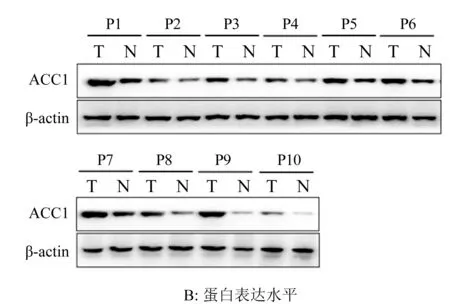
圖1 卵巢癌患者癌與癌旁組織中ACC1的表達
2.2 siRNA下調對SKOV3細胞中ACC1表達和細胞增殖的影響 為研究ACC1在卵巢癌細胞增殖中的調控作用,首先合成了2條靶向ACC1不同位置的siRNA干涉片段(分別命名為si-ACC1#1與si-ACC1#2),通過用qRT-PCR與Western Blot分析二者干涉效率后發現(圖2A與2B):轉染2條siRNA(si-ACC1#1與si-ACC1#2)均可顯著下調SKOV3細胞中ACC1的表達(P<0.01),且二者效率相當。

圖2 siRNA下調SKOV3細胞中ACC1表達的qRT-PCR與Western Blot驗證
用MTS實驗分析下調ACC1對卵巢癌細胞SKOV3增殖的影響,結果表明下調ACC1表達后卵巢癌細胞的增殖被顯著抑制(P<0.01;圖3)。
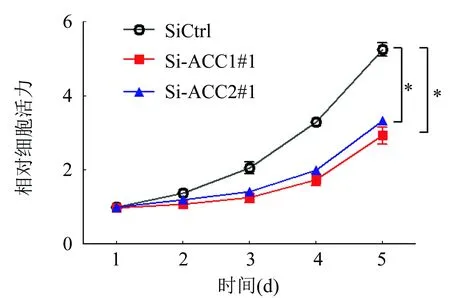
圖3 MTS實驗分析下調ACC1對SKOV3細胞增殖的影響
利用EDU實驗分析下調ACC1對卵巢癌細胞SKOV3增殖的影響,結果與MTS相一致,下調ACC1表達后,卵巢癌細胞的增殖被顯著抑制(P<0.01;圖4)。
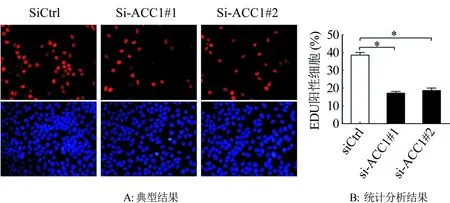
圖4 EDU實驗分析下調ACC1對SKOV3細胞增殖的影響
2.3 siRNA下調對SKOV3細胞凋亡的影響 進一步又利用流式細胞術分析下調ACC1對卵巢癌細胞凋亡的影響,結果表明下調ACC1表達后,卵巢癌細胞的凋亡未發生顯著改變(P>0.05;圖5)。
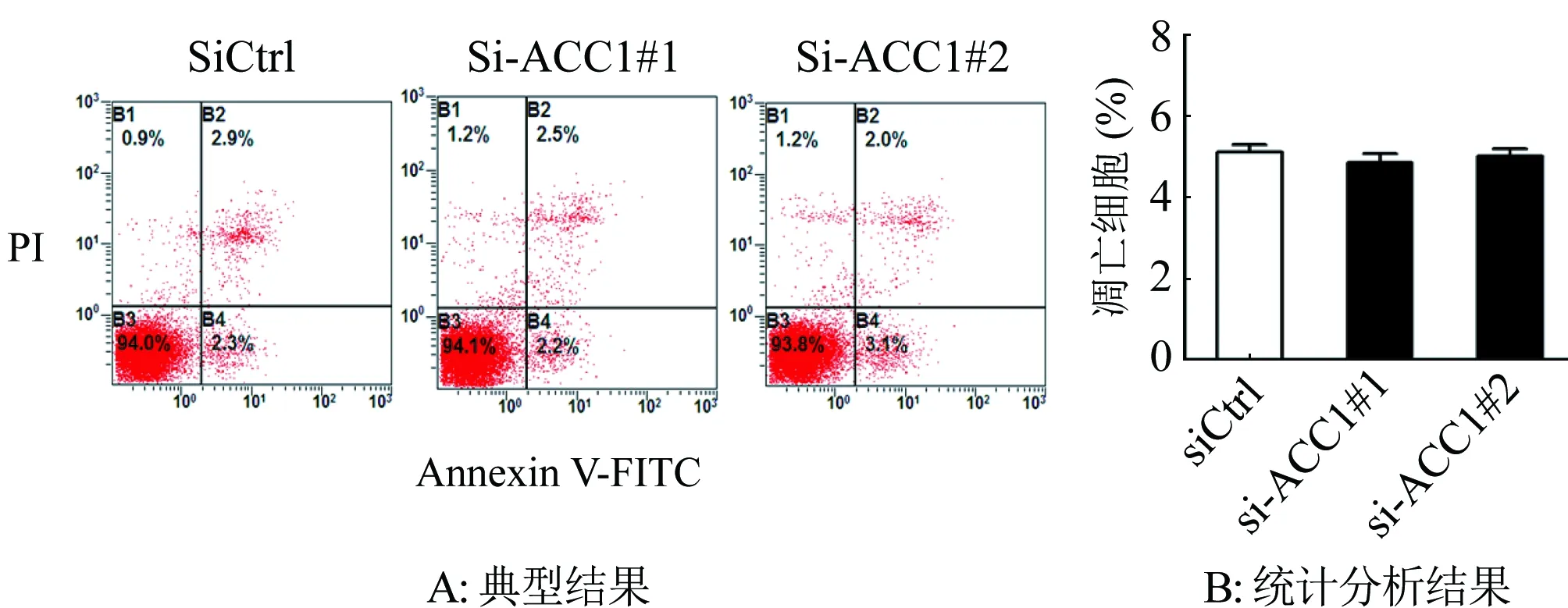
圖5 流式細胞術分析下調ACC1對SKOV3細胞凋亡的影響
3 討論
Cell雜志在2011發表權威綜述文章,總結了腫瘤細胞的十大最主要特征,其中包括了腫瘤細胞代謝的重編程[18]。腫瘤細胞代謝重編程的主要表現包括糖酵解、脂肪酸從頭合成和谷氨酰胺代謝的異常活躍[19]。而其中,與糖酵解和谷氨酰胺代謝相比,腫瘤細胞脂肪酸代謝受到的關注相對較少。但目前研究已證實,脂肪酸從頭合成增強是大多數類型腫瘤細胞所具有的共同代謝特點,在腫瘤發生發展中發揮了重要的促進作用[17,20]。腫瘤細胞通過大量合成游離脂肪酸,為其快速分裂增殖所需的生物膜等結構性成份的合成提供脂類原料[9]。此外,多種脂肪酸衍生物,如溶血磷脂酸[21-22]和人前列腺素[23]等,均是細胞內重要的參與生存、增殖和轉移調控的信號分子[11]。
ACC1是調控脂肪酸合成第一步反應的關鍵酶,催化乙酰輔酶A羧化生成丙二酰輔酶A。近年來,多個研究證實,ACC1在肝癌、肺癌、乳腺癌與胰腺癌中表達均異常升高。Wang等[12]研究發現肝癌細胞中ACC1表達顯著上調,ACC1可通過促進肝癌細胞中脂含量儲存,而促進能量應激時腫瘤細胞的生存。Svensson等[13]在肺癌中研究也證實,抑制ACC1可通過抑制脂肪酸合成而抑制非小細胞肺癌的生長。Chajes等[14]在乳腺癌中研究也發現,用siRNA下調ACC1可降低細胞內脂肪酸含量,并能誘導乳腺癌細胞發生凋亡。同樣,Petrova等[15]研究發現,ACC1在胰腺癌中顯著高表達,通過小分子抑制劑抑制ACC1活性后,腫瘤細胞中蛋白質的脂化與脂相關信號均被顯著抑制,并最終使腫瘤細胞的增殖變慢。此外,Corominas-Faja等[24]研究發現,用小分子化合物抑制ACC1活性后,乳腺癌干細胞的自我更新被顯著抑制。
以往研究表明,卵巢癌中脂類信號分子,尤其是溶血磷脂酸的水平顯著升高并調控腫瘤多個惡性生物學過程[25],表明脂代謝異常與卵巢癌發生發展密切相關。然而目前,卵巢癌組織細胞中ACC1等調控脂肪酸代謝關鍵酶的表達與作用尚不十分清楚。本研究在卵巢癌中發現:ACC1在卵巢癌中表達顯著上調;下調ACC1可顯著抑制卵巢癌細胞的增殖,而不影響其凋亡。我們的研究結果進一步表明卵巢癌細胞中脂肪酸合成活性異常增強,并提示通過靶向ACC1分子抑制脂肪酸合成可能是潛在的卵巢癌治療分子靶標。但本研究尚缺乏ACC1對卵巢癌生長與凋亡調控作用的體內實驗證據。此外,導致卵巢癌細胞中ACC1表達上調及ACC1調控細胞增殖與凋亡的分子機制仍有待進一步闡明。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發明與創新(2016年38期)2016-08-22 03:02:52
