武夷山中亞熱帶常綠闊葉林土壤氮礦化的季節動態
2019-08-22 02:08:26葛曉敏陳水飛周旭徐輝胡亞萍江寶興丁暉
生態環境學報 2019年7期
葛曉敏,陳水飛,周旭,徐輝,胡亞萍,江寶興,丁暉*
1. 生態環境部南京環境科學研究所自然保護與生物多樣性研究中心/國家環境保護武夷山生態環境科學觀測研究站/武夷山生物多樣性綜合觀測站/國家環境保護生物安全重點實驗室,江蘇 南京 210042;2. 福建省武夷山生物研究所,福建 武夷山 354300
土壤中的氮絕大部分以有機態的形式存在,約占全氮量的92%-98%。然而大多數的有機態氮不能被植物直接吸收利用,必須通過微生物礦化作用將有機氮轉化為無機氮才可以被植物吸收利用。氮礦化過程是土壤提供氮素養分的重要過程,礦化速率決定了植物生長對土壤氮素的可利用性(Koblerg et al.,1997)。氮礦化也是森林生態系統氮素循環最重要的過程之一,其對于揭示生態系統功能、生物地球化學循環本質具有重要意義。
土壤氮礦化過程受許多因素綜合作用的影響,環境因子(如土壤溫濕度、pH等)影響土壤生物的組成和結構,對氮礦化具有重要的調節作用。土壤溫度是影響微生物活性一個重要驅動因子,其在調節土壤氮礦化和氮的有效性方面起著重要的作用(Kabba et al.,2004;Wang et al.,2006)。大量室內培養或野外研究發現凈氮礦化速率隨溫度的升高而增加(Dalias et al.,2002;Yin et al.,2012),但也有研究表明,在5-15 ℃范圍內,氮礦化速率與溫度的相關性較弱(Lang et al.,2010;高俊琴等,2008)。多數研究認為,在一定的濕度范圍內,土壤氮礦化速率與土壤濕度呈顯著的線性正相關關系,但是,當土壤含水量超過一定范圍時,氮礦化速率會明顯下降(李貴才等,2001;周才平等,2003)。此外,海拔梯度和季節變化引起的土壤溫濕度等生態因子的變化,也有可能影響土壤氮礦化速率。有研究發現,森林土壤凈氮礦化速率隨海拔升高而增加(Knoepp et al.,1998;Bonito et al.,2003),也有研究得出相反的結果(劉金玲等,2012;劉琪璟等,2009),還有研究認為,土壤凈氮礦化速率與海拔無直接關系,其主要由土壤C/N所決定(李菊梅等,2003)。陳書信等(2014)的研究表明,蘇北沿海地區楊樹林、楊農復合、草地和農田等 4種不同土地利用方式的土壤氮礦化速率季節變化顯著,最大值出現在春季。孫磊(2012)研究則表明,香樟(Cinnamomum camphora L.)、楊樹(Populus L.)和廣玉蘭(Magnolia grandiflora L.)3種典型生態防護林帶的土壤氮硝化速率和氮礦化速率在夏季達到最大值;張文文等(2016)在蘇北沿海地區的研究也表明楊樹人工林、農田、楊農復合林 3種土地利用類型的土壤凈氮礦化速率在夏季或秋季最大。
武夷山是中國重要生態功能區和生物多樣性保護優先區域,武夷山保存了世界同緯度帶最完整、最典型、面積最大的中亞熱帶原生性森林生態系統,其水源涵養、土壤保持及維系生物多樣性等重要生態功能對保障華東地區乃至全國的生態安全具有舉足輕重的作用。目前,對武夷山地區中亞熱帶常綠闊葉林的研究主要集中在物種組成與群落結構特征、土壤微生物群落特征、土壤呼吸以及土壤有機碳礦化特征等方面(吳則焰等,2014;金裕華,2012;丁暉等,2015;施政,2007;周焱等,2008),對該地區土壤氮礦化過程研究不足。本研究采用原位土柱-離子交換樹脂包法,通過野外原位試驗研究了武夷山常綠闊葉林生態系統土壤氮礦化過程和規律,分析了不同坡位土壤氮礦化過程以及與之相關的土壤溫度和濕度對土壤氮礦化的影響,為武夷山常綠闊葉林生態系統氮素養分管理和生物多樣性保護提供理論依據。
1 研究區概況
研究區位于武夷山國家自然保護區,研究地點處于武夷山市星村鎮四新村附近,地理位置為27°35′24.23″N,117°45′55.43″E。該區域屬中亞熱帶季風濕潤氣候,年平均氣溫17.0-18.4 ℃,平均相對濕度75%-84%,年降水量1800 mm,年平均日照時數1910.2 h,無霜期227-246 d。紅壤為境內分布最廣的一種地帶性土壤。常綠闊葉林是武夷山的地帶性森林植被,分布面積很大,多分布在海拔350-1400 m之間。
研究樣地海拔450-580 m,呈長方形,投影面積 9.6 hm-2(400 m×240 m),樣地坡度 10-50°。喬木層主要有米櫧(Castanopsis carlesii)、毛錐(Castanopsis fordii)、甜櫧(Castanopsis eyrei)等常綠闊葉樹種;灌木層主要有赤楠(Syzygium buxifolium)、微毛柃(Eurya hebeclados)、杜莖山(Maesa japonica)等;草本層主要有中華里白(Hicriopteris chinensis)、狗脊(Woodwardia japonica)、芒萁(Dicranopteris dichotoma)和蕨狀薹草(Carex filicina)等(丁暉等,2015)。
2 材料與方法
2.1 試驗設計與采樣
2017年 1月,選擇研究區南坡的上坡(U)、中坡(M)和下坡(L)為小樣地,每個坡位小樣地隨機設置3個5 m×5 m的樣方作為重復。3個坡位不同土層(0-10 cm土層和10-20 cm土層)的土壤基本理化性質見表1。
分別于2017年1月17日-4月19日(春季)、2017年4月20日-7月18日(夏季)、2017年7月19日-10月25日(秋季)、2017年10月26日-2018年1月16日(冬季)采用原位土柱-離子交換樹脂包法(ISC-IERB)測定原位土壤氮礦化速率。從2017年1月17日開始,在研究樣地的樣方內設置土壤原位氮礦化裝置,即將長10 cm,外徑5 cm,內徑4.6 cm的PVC管用環刀托和鐵錘將其分別打入不同的土層中,取出 PVC管,將裝有離子交換樹脂和石英砂的長3 cm、外徑5 cm、內徑4.6 cm的PVC管緊連在土柱PVC管上,將其原位埋入土壤中。每個離子交換樹脂包中含Amberjet陰陽離子交換樹脂22 g∶(陰陽=2∶1)。每個樣方每個土層各設置4個PVC管。每次取回埋置的PVC管后,在鄰近樣地上再埋置一批新的 PVC管,同時取回一定量的土壤樣品,作為土壤氮礦化的初始值,于實驗室內分析土壤和樹脂中的銨態氮、硝態氮含量以及土壤中的微生物生物量氮含量。
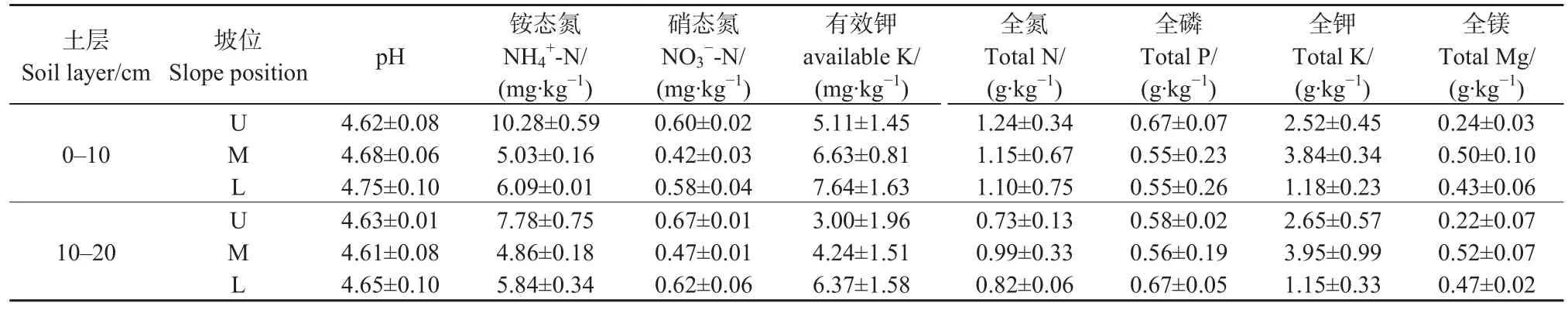
表1 試驗樣地土壤基本理化性質Table 1 Basic soil physical and chemical properties of soil in the plot of an evergreen broad-leaved forest
此外,自2017年4月開始,在每個坡位上安裝1臺Em50土壤水分監測系統(DECAGON,美國),每半個小時自動監測、記錄不同坡位不同土層土壤溫度和濕度。
2.2 測定方法
土壤含水量測定采用 105 ℃烘干稱重法;pH值的測定采用電位法(水∶土=2.5∶1);土壤容重的測定采用環刀法;采用2 mol·L-1KCl浸提土壤樣品,銨態氮含量測定采用靛酚藍比色法,硝態氮含量測定采用紫外分光光度法;土壤有效鉀采用1 mol·L-1乙酸銨浸提-原子吸收分光光度計測定(Solaar Unicam 969 AAS,USA);土壤經風干后,過 0.25 mm篩,使用H2SO4-HClO4消煮,全量N含量采用靛酚藍比色法測定,全量P含量采用碳酸氫鈉-鉬銻抗比色法測定,全量K和Mg含量由原子吸收分光光度計測定(Solaar Unicam 969 AAS,USA)(中國科學院南京土壤研究所,1978;中華人民共和國林業部科技司編,1991;魯如坤,1999);土壤微生物生物量氮(MBN)含量采用氯仿薰蒸-浸提法提取,靛酚藍比色法測定(吳金水等,2006)。
2.3 計算方法

式中,ω(mineral N)為礦質氮質量分數,mg·kg-1;Ramm為銨化速率,mg·kg-1·d-1;Rnit為硝化速率,mg·kg-1·d-1;Rmin為凈氮礦化 速率,mg·kg-1·d-1;ω(Pre NH4+-N)為培養前 NH4+-N 質量分數;ω(Post NH4+-N)為培養后NH4+-N質量分數;ω(Pre NO3--N)為培養前NO3--N質量分數;ω(Post NO3--N)為培養后NO3--N質量分數;t為培養天數。納入計算的NH4+-N和NO3--N包含土壤和樹脂包中樹脂吸附的NH4+-N和NO3--N。
2.4 數據處理
運用 Microsoft Excel 2007進行數據整理及繪圖,SPSS 18.0進行數據處理,采用兩因素方差分析(Two-way ANOVA)檢驗坡位和季節及其交互作用對土壤銨態氮、硝態氮、氨化速率、硝化速率、凈氮礦化速率和土壤微生物生物量氮的影響,并采用單因素方差分析(One-way ANOVA)和最小顯著差異法(LSD)檢驗不同數據間的差異顯著性,采用Pearson相關性分析檢驗土壤溫濕度與土壤礦質氮、微生物生物量氮含量和礦化速率間的相關關系。
3 結果與分析
3.1 不同坡位土壤溫度和濕度的動態變化
由圖1可知,武夷山地區中亞熱帶常綠闊葉林土壤溫度總體上呈單峰變化。不同坡位不同土層土壤月平均溫度表現為8月最高,2月最低。上坡、中坡、下坡0-10 cm土層土壤年平均溫度分別為16.9、17.1、17.3 ℃;上坡、中坡、下坡10-20 cm土層土壤年平均溫度分別為17.1、17.3、17.4 ℃,呈現坡位越高,土壤溫度越低的特征。不同土層土壤年平均溫度表現為0-10 cm土層(17.1℃)<10-20 cm土層(17.3 ℃)。
不同坡位不同土層土壤月平均濕度表現為6月最大,10月或11月最小。上坡、中坡、下坡的0-10 cm土層土壤年平均濕度分別為0.266、0.232、0.214 m3·m-3,呈現土壤濕度隨坡位上升而增加的變化趨勢;上坡、中坡、下坡10-20 cm土層土壤年平均濕度分別為 0.221、0.225、0.226 m3·m-3,表現為土壤濕度隨坡位上升而降低。不同土層土壤年平均濕度表現為0-10 cm土層(0.237 m3·m-3)>10-20 cm 土層(0.224 m3·m-3)。
3.2 不同坡位土壤銨態氮、硝態氮、礦質氮含量和凈氮礦化量的季節變化
研究期間,各坡位的土壤銨態氮和氮含量含量存在明顯的季節差異(圖2,表2)。總體上,各坡位夏季和秋季的土壤銨態氮含量(12.02-81.78 mg·kg-1)高于春季和冬季(4.86-44.89 mg·kg-1),且隨著坡位上升土壤銨態氮含量逐漸增大;就不同土層而言,0-10 cm土層土壤銨態氮含量(6.09-81.78 mg·kg-1)高于10-20 cm土層的含量(4.86-52.37 mg·kg-1)。上坡0-10 cm土層,秋季的土壤硝態氮含量顯著高于其他季節(P<0.05);不同坡位土壤硝態氮含量大致表現為上坡和下坡較高,中坡較低。土壤礦質氮(銨態氮與硝態氮之和)含量的變化趨勢與土壤銨態氮基本一致,表明銨態氮是該地區土壤礦質氮的主要組成。坡位與采樣季節的交互作用對土壤銨態氮和礦質氮含量有顯著影響(P<0.05)。
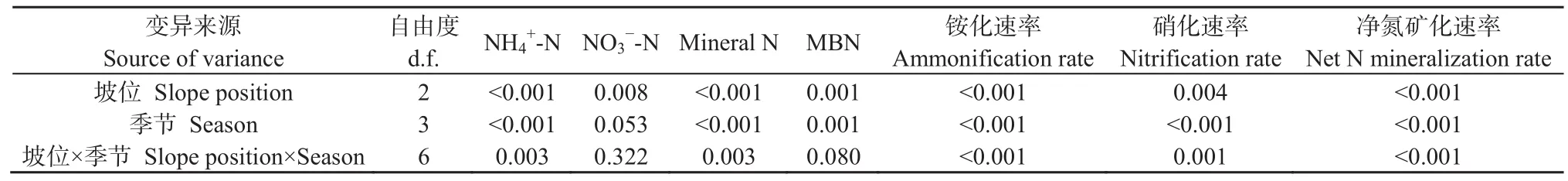
表2 武夷山常綠闊葉林不同坡位和季節對土壤礦質氮含量、礦化速率和MBN含量的方差分析Table 2 Analysis of variance (P values) for soil mineral nitrogen contents, nitrogen mineralization rates and MBN content to slope position and season
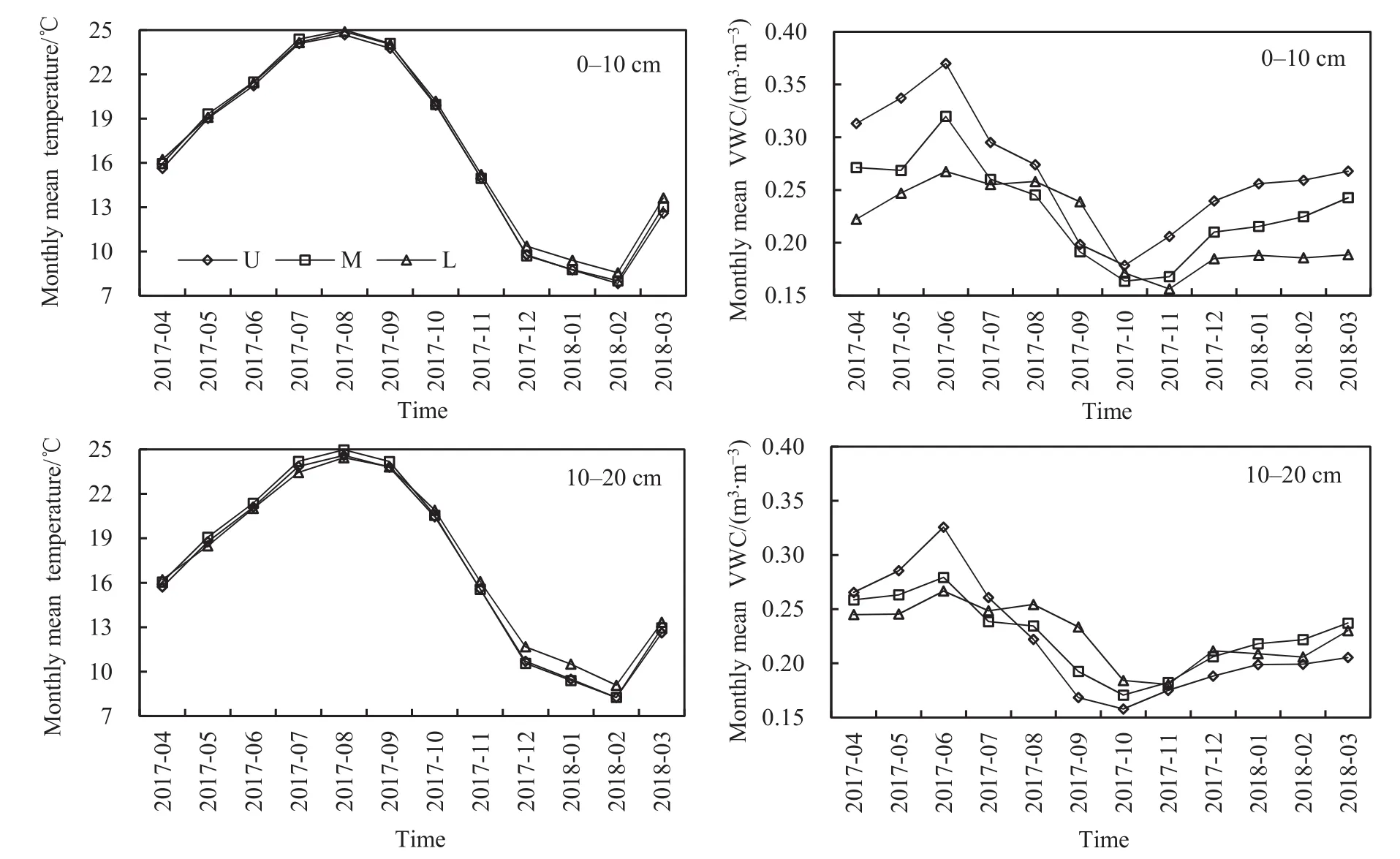
圖1 不同坡位不同土層土壤溫度和濕度動態變化Fig. 1 Temporal fluctuations of soil temperature and moisture in different slope positions of different soil layers
如表3所示,不同土層不同坡位的土壤凈氮礦化量存在季節差異。上坡土壤凈氮礦化量在 4-7月間最大,0-10 cm和10-20 cm土層分別為56.3 kg·hm-2和 50.3 kg·hm-2;下坡 7-10 月土壤凈氮礦化量顯著高于其他培養時期(P<0.05)。0-10 cm土層土壤年凈氮礦化量為 42.4-131.4 kg·hm-2·a-1,高于 10-20 cm 土層的 44.0-93.0 kg·hm-2·a-1;不同土層的年凈氮礦化量表現為上坡>中坡或下坡。
3.3 不同坡位土壤微生物生物量氮含量的季節變化
不同坡位不同季節間的土壤MBN含量存在一定差異(圖3,表2)。2個土層土壤MBN含量總體趨勢表現為夏季和秋季(7.49-73.60 mg·kg-1)較高,春季(11.69-22.47 mg·kg-1)次之,冬季(5.50-14.21 mg·kg-1)最低;就不同坡位而言,總體上表現為上坡高于中坡和下坡。其中0-10 cm土層,夏、秋兩季上坡土壤的MBN含量顯著高于中坡或下坡(P<0.05)。
3.4 不同坡位土壤銨化速率、硝化速率和凈氮礦化速率的季節變化
如圖4所示,不同季節不同坡位的土壤銨化速率和硝化速率存在一定差異。在0-10 cm土層中,總體上,2017年4-7月和7-10月的土壤銨化速率高于其他培養時期。銨化速率的坡位變化中,上坡高于中坡和下坡;而在10-20 cm土層中,4-7月和7-10月上坡土壤的銨化速率顯著高于中坡和下坡(P<0.05)。0-10 cm土層土壤銨化速率(-0.13-1.13 mg·kg-1·d-1)總體上大于 10-20 cm 土層(-0.10-0.74 mg·kg-1·d-1)。2017 年 4-7 月間的土壤硝化速率(0.013-0.025 mg·kg-1·d-1)顯著高于其他培養時期(P<0.05)。土壤凈氮礦化速率的總體變化趨勢與銨化速率基本一致。坡位與采樣季節的交互作用對土壤銨化速率、硝化速率和凈氮礦化速率均有顯著影響(P<0.05)(表 2)。
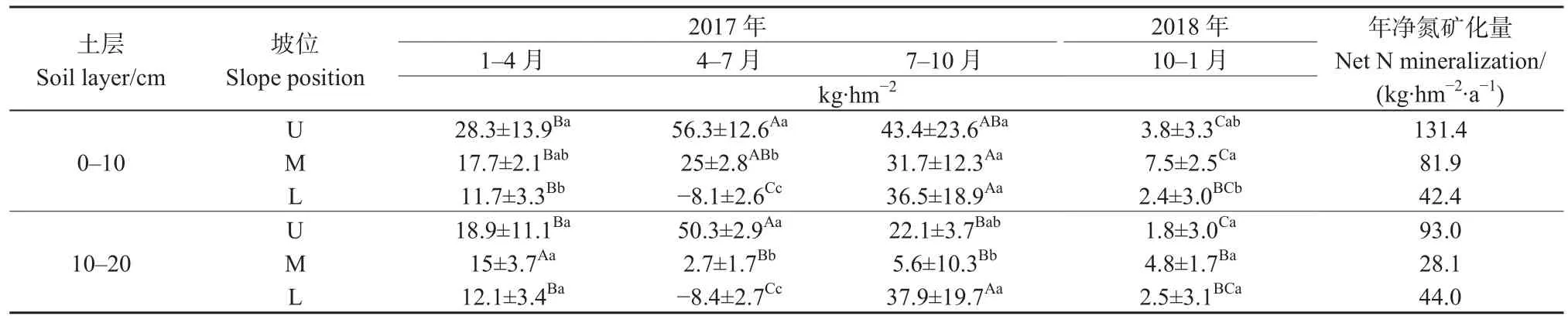
表3 不同土層和坡位土壤凈氮礦化量Table 3 Soil in situ net N mineralization accumulation in different slope positions of various soil depths (0-10 and 10-20 cm) at different sampling times in an evergreen broad-leaved forest
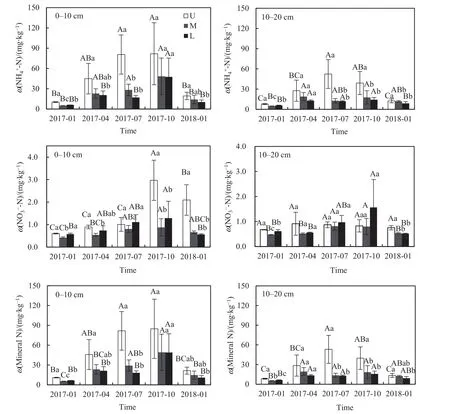
圖2 不同坡位不同土層土壤銨態氮、硝態氮和礦質氮含量季節變化Fig. 2 Seasonal fluctuations of soil NH4+-N, NO3--N and mineral N contents in different slope positions of various soil depths (0-10 cm and 10-20 cm)
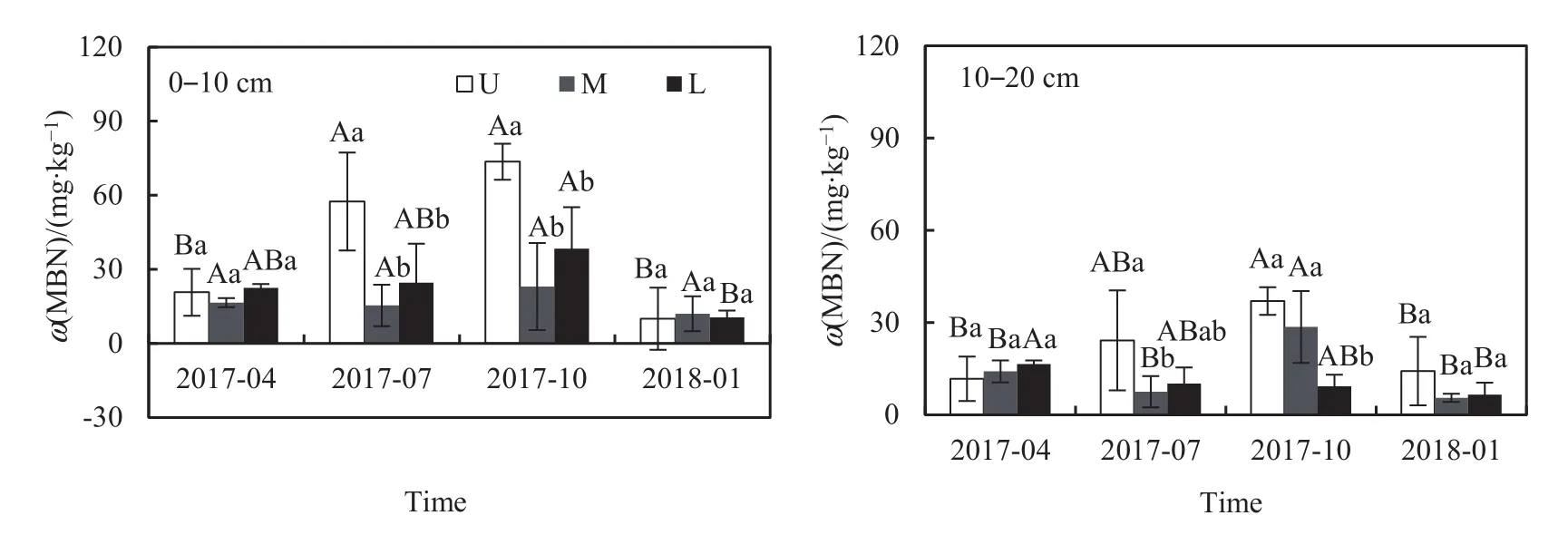
圖3 不同坡位不同土層土壤MBN含量季節變化Fig. 3 Seasonal fluctuations of soil MBN content in different slope positions of various soil depths (0-10 and 10-20 cm)
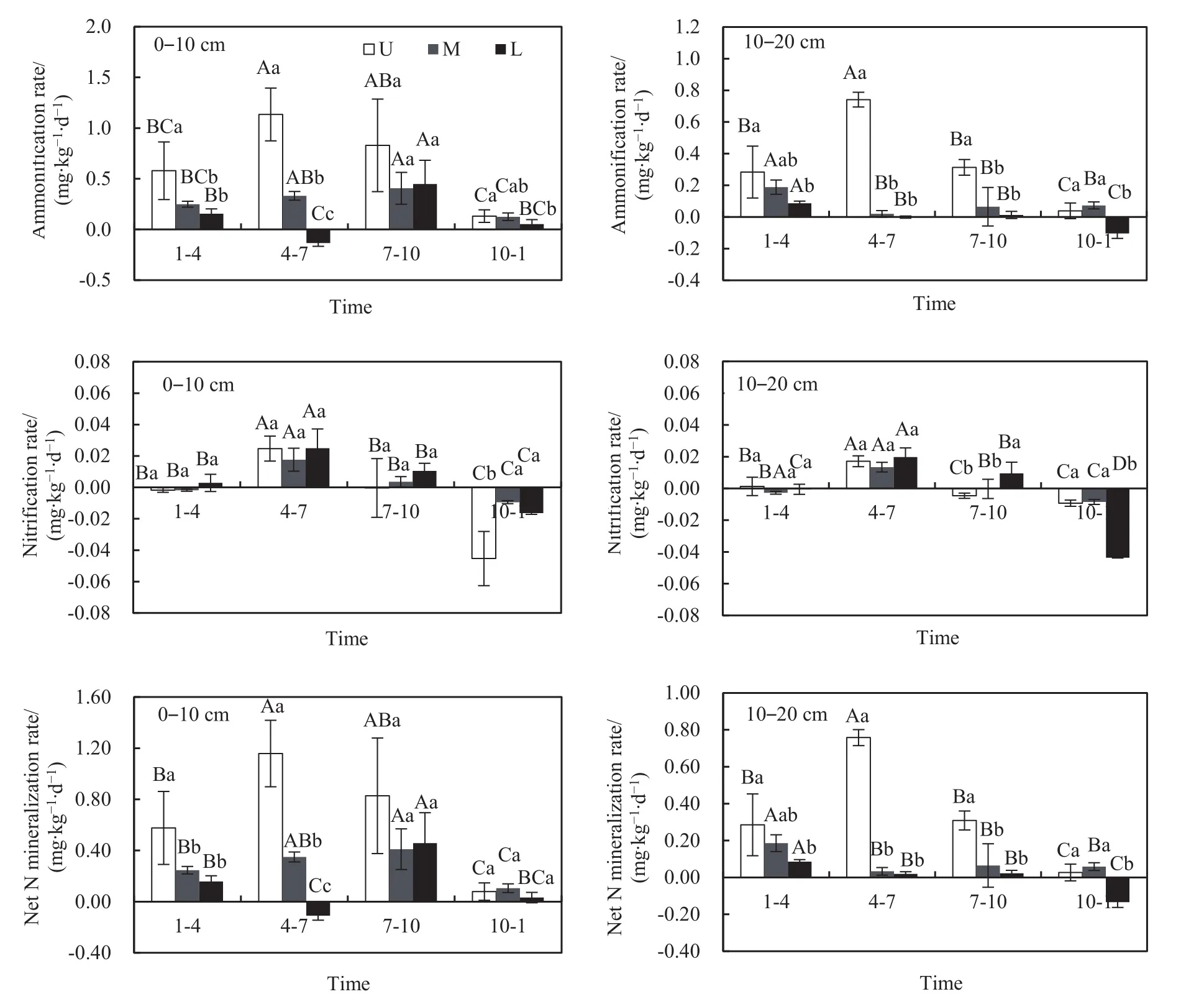
圖4 不同坡位不同土層土壤銨化、硝化和凈氮礦化速率的季節變化Fig. 4 Seasonal fluctuations of soil ammonification rate, nitrification rate and net N mineralization rate in different slope positions of various soil depths(0-10 cm and 10-20 cm)
3.5 土壤溫濕度與土壤礦質氮、微生物生物量氮含量和礦化速率的相關分析
如表 4所示,土壤溫度與銨態氮、礦質氮和MBN含量呈顯著正線性相關(P<0.05),與硝化速率呈極顯著正線性相關(P<0.01);土壤濕度與銨化速率、硝化速率和凈N礦化速率呈極顯著正線性相關(P<0.01)。
4 討論
4.1 季節、坡位和土層因素對土壤礦質氮含量及礦化速率的影響
氮礦化是森林生態系統氮循環過程中的重要一環,氮礦化速率往往決定了植物生長所能吸收利用的氮含量(林貴剛等,2012;王光軍等,2009)。本研究結果顯示,武夷山地區常綠闊葉林大樣地土壤銨態氮是土壤礦質氮的主要組成,土壤礦化過程中銨化作用起決定性作用。這一結果與李巍等(2015)和徐憲根(2009)等在武夷山地區的研究結果一致,也與前人在其他地區的研究結果一致(莫江明等,1997;孟盈等,2001;陳伏生等,2004)。武夷山地區礦質氮含量出現銨態氮占主導的現象,主要與土壤的pH值有關,中性或堿性土壤最適宜硝化作用的進行,而經過前期調查發現武夷山地區土壤的pH值在4.4-4.9之間,土壤呈酸性,不利于該地區土壤硝化作用的進行。也有研究認為出現這一情況是由于森林生態系統根系層pH值低并富含單寧,因而硝化細菌數量少,硝化速率低,而這一現象也可能是生態系統保存氮的一種機制(沙麗清等,2000)。
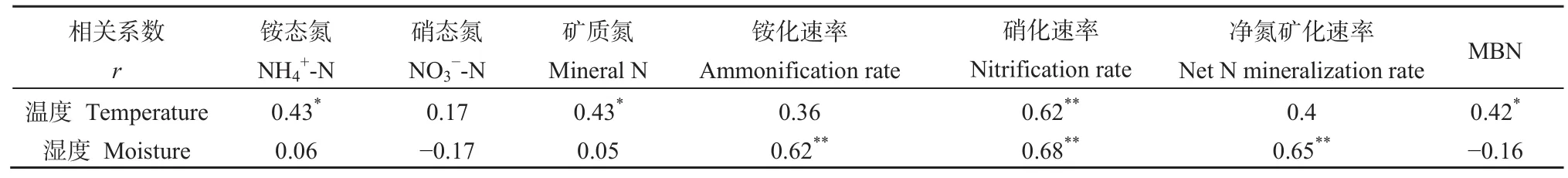
表4 土壤溫濕度與土壤礦質氮、微生物生物量氮含量和礦化速率的相關分析Table 4 Correlation coefficient (r) between soil temperature or moisture and soil mineral N, MBN contents, or N mineralization rates(included 0-10 and 10-20 cm soil layer)
坡位與海拔密切相關,一般坡位越高,海拔越高。本研究結果顯示,武夷山地區常綠闊葉林生態系統土壤銨態氮和礦質氮含量、銨化速率和凈氮礦化速率,總體上隨著坡位升高而增大。這一結果與殷睿等(2013)的研究結果一致,Bonito et al.(2003)的研究也發現美國南部高海拔硬木林土壤中氮的礦化速率比低海拔處高。高海拔地塊土壤銨態氮和銨化速率較高,可能是因為海拔越高土壤溫度越低,土壤溫度較低時銨態氮不易轉化成硝態氮(Cookson et al.,2007),致使銨態氮積累,表現為銨態氮含量較大。且隨著坡位的上升,研究樣地土壤pH值,尤其是上層土壤的pH值越低(表1),較低的pH值也會在一定程度上抑制硝化細菌的繁殖和生長,弱化硝化作用,硝化速率和硝態氮含量相對較低。
土壤有機氮的礦化主要受控于土壤微生物,而微生物的種類、數量與活性在一定程度上又受土壤溫度和濕度的影響,所以土壤溫度與濕度是影響土壤氮礦化的2個重要外部因子(葛曉敏等,2015),然而土壤溫濕度與不同季節的氣候條件密切相關。本研究發現季節性的氣候變化造成的土壤溫度和濕度變化是影響武夷山地區常綠闊葉林生態系統土壤原位氮礦化的主要因素之一。隨著季節變化,土壤礦質氮含量的高低與土壤生物活性、土壤有效氮的淋失及植物的吸收等相關。本研究結果顯示土壤溫度最大值出現在8月,土壤濕度最大值出現在6月,二者的最低值分別在2月和11月,即土壤溫度最大值出現在夏季,最低值出現在冬季。總體上,土壤銨態氮、礦質氮含量以及銨化速率、硝化速率和凈氮礦化速率均表現為夏季最高,秋季次之,冬季最低。這一結果與Yan et al.(2015)在蘇北楊樹人工林內的研究結果一致。這主要是因為在夏、秋季,武夷山地區進入雨季,降水量增大,氣溫逐漸升高,土壤溫濕度較高,有利于土壤微生物及土壤酶活性的提高,進而加速有機氮的礦化,而在深秋入冬季節,該地區進入干季,氣溫下降,降雨減少,土壤微生物活性逐漸下降,致使土壤的礦化速率減弱。其他地區研究結果則與武夷山地區有所不同,如內蒙古鄂爾多斯地區研究結果顯示表土層(0-10 cm)銨態氮和硝態氮的最大值均出現在春季(史作民等,2004),黑龍江涼水自然保護區各林型 A0層土壤銨態氮和硝態氮的含量為春季高于夏季,峰值出現在秋季(許翠清等,2008)。可見,由于研究地點的不同,土壤理化性質、生物特性以及氣候特征都存在差異,從而使不同研究區所得出的結論有所不同。
土壤濕度也是影響土壤硝態氮含量的主要因素之一。本研究發現,該地區土壤銨態氮和總礦質氮含量與土壤濕度呈正線性相關關系(表4),而硝態氮含量則與土壤濕度呈負線性相關關系,這可能是因為過高的土壤濕度或含水量影響土壤的通氣結構,不利于土壤硝化細菌的生長。此外,較高的土壤含水量會導致土壤反硝化作用增強,造成大量硝態氮以氧化亞氮(N2O)形式從土壤逸失到大氣中,進一步造成硝態氮的損失。最后,硝態氮帶負電荷,極易在該地區大量降水時隨降水淋溶流失,導致硝態氮含量較低,而銨態氮為可交換態陽離子,易被帶負電荷的土壤粘粒和膠體所吸附,不易從土壤中淋溶流失。總之,該地區土壤總無機氮庫主要由銨態氮組成,可能是氣候、土壤和植被等多因子綜合作用的結果。
土層差異對武夷山地區常綠闊葉林土壤氮礦化也存在一定影響。本研究結果表明,不同土層間的土壤礦質氮含量、氮礦化速率差異明顯。隨著土層加深,上層土壤礦質氮含量均大于下層,這與徐憲根(2009)在武夷山的研究結果,韓玉萍等(1998)和楊萬勤等(2001)在縉云山的研究結果一致。上層土壤(0-10 cm)的礦化速率高于下層土壤(10-20 cm),這主要是因為上層土壤的有機質含量高于下層,微生物分解礦化的底物豐富(Arunachalam et al.,1998),其次,上層土壤的理化性質優于下層土壤,更利于微生物的生長繁殖及生物活性的提高。然而,上層土壤相較于下層土壤,其對氣候變化的響應更加敏感,氣候波動對上層土壤N礦化的影響也高于下層土壤。
4.2 季節、坡位和土層因素對土壤 MBN含量的影響
土壤微生物在森林生態系統養分循環中具有重要作用(Singh et al.,1989)。微生物的種群結構和數量與土壤理化性質密切相關(Xue et al.,2006)。而微生物生物量氮(MBN)是活性氮庫中最重要的氮素形態之一,是土壤有效氮的來源之一,其輕微的變化可以影響土壤氮素循環過程和森林土壤的供氮能力。本研究結果表明,上層(0-10 cm)土壤中的微生物生物量氮顯著高于下層(10-20 cm)土壤,這主要是因為不同土層氧氣有效性和土壤有機質含量不同,進而影響土壤微生物的生長速率,致使微生物生物量不同(Cao et al.,2008)。
坡位差異對土壤MBN含量也有一定影響。研究結果表明,不同土層不同季節的土壤MBN含量總體表現為上坡高于中坡和下坡,即海拔較高的樣地土壤MBN含量高于海拔較低的樣地。這一結果與殷睿等(2013)在川西亞高山不同海拔森林發現不同采樣時期微生物生物量氮基本隨海拔降低呈升高的趨勢的結果有所不同。前人的研究認為海拔越低,土壤溫度越高,微生物活性越強,則微生物生物量氮越高(Li et al.,2004)。出現這一差異可能是因為該地區研究樣地不同坡位間土壤溫差較小,溫度變化不是影響土壤MBN含量的主要因素,而土壤有機質含量、植物根系分布等綜合因素導致的土壤微生物和土壤酶活性的差異可能是主導不同坡位間土壤MBN含量差異的主要因素。
研究結果顯示,武夷山地區常綠闊葉林生態系統土壤MBN含量在秋季和夏季較高,春季次之,冬季最低;MBN含量的季節動態變化規律與該地區土壤銨態氮、礦質氮含量隨季節的動態變化基本一致。這主要是因為秋季和夏季是武夷山地區的雨季,高溫高濕的土壤環境利于土壤微生物的生長繁殖,相反地,春季和冬季是該地區的干季,不利于土壤微生物的繁殖與生長,而且MBN作為活性氮庫中最重要的氮素形態之一,在干季時可作為有效氮源,為植物的生長提供生長所需氮素,進而導致其在干季時含量較低。然而,不同研究者在不同研究地點得出的結論也不盡相同。Piao et al.(2000)在中國西南喀斯特地區的研究發現土壤微生物生物量碳在冬季最高,夏季最低;而 Amold et al.(1999)在云杉(Picea asperata Mast.)和冷杉(Abies fabri (Mast.)Craib)混交林中發現微生物生物量及活性隨土壤溫度、濕度和基質有效性的季節模式的變化而變化,土壤微生物生物量氮一般在春秋季最高,夏冬季最低。出現這種微生物生物量不同的季節變化模式可能主要是因為不同生態系統土壤微生物組成結構不同,而不同的微生物種類對季節變化的響應有一定的差異。
氮是植物生長和發育所需的大量營養元素,是調節森林生態系統生產量、結構和功能的關鍵元素之一。本研究基本探明了武夷山常綠闊葉林生態系統土壤有機氮礦化的坡位效應和季節動態變化的基本規律,在今后的研究工作中,將持續開展該生態系統的氣象、水文、土壤性質等環境因子的監測,以及植被多樣性的長期觀測,探討環境因子對武夷山常綠闊林生態系統生物多樣性的影響,為該地區生物多樣性保護研究與管理提供一定的理論依據。
5 結論
(1)武夷山地區中亞熱帶常綠闊葉林土壤溫度總體上呈單峰變化,坡位越高,土壤溫度越低。
(2)土壤銨態氮含量占總礦質氮含量 90%以上,表明土壤礦化過程中銨化作用起決定性作用。
(3)土壤銨態氮、礦質氮、MBN含量與銨化速率、凈氮礦化速率總體上在夏季或秋季較高,春季次之,冬季較小,表明土壤氮礦化存在明顯的季節效應。
(4)土壤銨態氮、礦質氮、年凈氮礦化量與銨化速率、凈氮礦化速率存在明顯的坡位效應,隨坡位的上升總體呈逐漸增大的趨勢。
(5)土壤溫度與硝化速率呈極顯著正線性相關(P<0.01),土壤濕度與銨化速率、硝化速率和凈N礦化速率呈極顯著正線性相關(P<0.01),表明土壤溫濕度是影響土壤氮礦化的關鍵因子。
