甜菜夜蛾幾丁質脫乙酰酶SeCDA7的表達與結合活性分析
2019-08-08 03:32:04張偉畢揚趙丹宗召莉郭巍
生物技術通報 2019年8期
張偉 畢揚 趙丹 宗召莉 郭巍
(1. 北京農學院植物科學技術學院,北京 102206;2. 中國農業科學院蔬菜花卉研究所,北京 100081;3. 河北農業大學植物保護學院,保定 071001)
幾丁質是昆蟲表皮和圍食膜的重要組成成分,幾丁質代謝隨昆蟲不同生長發育階段而變化,對昆蟲的正常生長發育至關重要[1],參與幾丁質代謝的酶是殺蟲劑破壞昆蟲生長機制的潛在靶標。幾丁質脫乙酰酶(Chitin deacetylase,CDA)是昆蟲幾丁質代謝酶系中的重要組成,可催化幾丁質脫乙酰化形成殼聚糖[2]。目前,對多種昆蟲CDA蛋白功能研究表明,昆蟲CDA類蛋白與昆蟲蛻皮、羽化以及氣管延伸等方面有關,為昆蟲防治提供了有利依據。
昆 蟲CDA蛋 白 分 為CDAⅠ、CDAⅡ、CDAⅢ、CDAⅣ及CDAⅤ五大類[3]。CDAⅠ類蛋白包含幾丁質脫乙酰基酶催化區結構域(CDA)、幾丁質結合結構域(ChBD)以及低密度脂蛋白結合區(LDLa),CDAⅡ類蛋白結構域與第Ⅰ類的結構域一致,但是序列差異大,研究表明,赤擬谷盜(Tribolium castaneum)TcCDA1與TcCDA2與其蛻皮和羽化有關[4];飛 蝗(Locusta migratoria)LmCDA1、LmCDA2a 和LmCDA2b在昆蟲細胞中成功表達,并具有不同的幾丁質脫乙酰酶活性[5];CDAⅢ類蛋白和CDAⅣ類蛋白都具有CDA與ChBD結構域,不含有LDLa結構域,但是第Ⅲ類和第Ⅳ類CDA的結構域在CDA上的位置存在較大程度差異;CDAⅤ類蛋白只含有CDA結構域,不具有ChBD 與LDLa結構域,CDAⅤ類蛋白研究中,粉紋夜蛾CDA研究表明,TnPM-P42蛋白具有很強的幾丁質結合活性,與圍食膜結合緊密[6],蓓帶夜蛾CDA的研究結果推測CDA的作用可能是參與改變圍食膜中幾丁質的物理和化學性質,而這一生化活動對圍食膜蛋白的結合程度、圍食膜的完整性和孔隙率產生影響[7]。在黑腹果蠅中,克隆得到2個能編碼具有典型CDA功能域的CDA,通過氣管上幾丁質的脫乙酰基限制氣管延長[8]。赤擬谷盜第Ⅴ類CDA蛋白在中腸內特異性表達,利用RNA干擾技術對昆蟲CDA表達進行調控,干擾昆蟲正常生長發育和新陳代謝過程以及抵抗病原物的特性[9]。幾丁質代謝隨昆蟲不同生長發育階段而變化,對昆蟲的正常生長發育至關重要,因此,通過破壞幾丁質結構或幾丁質代謝的平衡來防治害蟲具有極大的發展潛力。
甜菜夜蛾Spodoptera exigua(Hübner),屬鱗翅目、夜蛾科,是以危害蔬菜為主的雜食性害蟲[10]。本研究克隆甜菜夜蛾幾丁質脫乙酰酶secda7基因,體外表達的SeCDA7蛋白進行幾丁質結合活性分析,利用實時定量PCR方法分析secda7基因轉錄水平在不同組織的表達,為明確深入探究昆蟲幾丁質脫乙酰酶的生理功能奠定理論基礎,為甜菜夜蛾生物防治提供材料。
1 材料與方法
1.1 材料
1.1.1 供試昆蟲與細胞培養 甜菜夜蛾為北京農學院植保系農藥實驗室利用人工氣候箱飼養;草地貪夜蛾昆蟲細胞系(Sf9)為北京農學院植保系農藥實驗室27℃培養。
1.1.2 實驗試劑、載體與菌株 質粒pMD19-T、pET-30a(帶有His標簽)和pFastBac HT A(帶有His標簽)以及E.coliDH5α、BL21、JM109和E.coliDH10BacTM菌株由本實驗室保存。
限制性內切酶、T4連接酶購自Promega公司;LA Taq酶、SYBR Premix Ex TaqTM購自Takara公司;RNA提取試劑盒購自QIAGEN公司;反轉錄試劑盒、質粒提取試劑盒購自Axygen公司;Mouse anti-6×His tag monoclonal antibody、Goat Anti-Mouse lgG(H+L)、NI-Agarose His標簽蛋白純化試劑盒購自康為試劑公司。
1.2 方法
1.2.1 甜菜夜蛾secda7基因的克隆及序列分析 根據甜菜夜蛾中腸轉錄組數據,設計secda7全長基因引物(表1)。提取甜菜夜蛾四齡幼蟲中腸RNA,以反轉錄cDNA為模板,以secda7 F和secda7 R為引物進行PCR,擴增甜菜夜蛾幾丁質脫乙酰酶cDNA序列片段。PCR產物經純化后連接pMD19-T,轉化DH5α感受態細胞,挑取單菌落進行雙酶切鑒定后序列測定,并對secda7基因進行生物信息學分析。利用DNAMAN軟件分析其開放閱讀框及其氨基酸序列,NCBI Blast進行功能域分析,使用SignalP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/) 進行信號肽預測,用NetNGlyc1.0(http://www.cbs.dtu.dk/services/NetNGlyc/)預測N-聯糖基化位點。
1.2.2secda7基因的原核表達 設計帶有酶切位點的引物ySecda7 F和ySecda7 R進行secda7基因的克隆(表1),將secda7與pET-30a原核表達載體使用T4連接酶進行連接反應,轉化BL21感受態細胞,挑取單菌落進行雙酶切鑒定。鑒定正確的陽性克隆于28℃培養時,分別加入不同濃度(0.25 mmol/L、0.5 mmol/L、0.75 mmol/L、1 mmol/L、1.25 mmol/L)的IPTG誘導蛋白表達,分別利用第一抗體小鼠抗6×His單克隆抗體(Mouse anti-6×His tag monoclonal antibody)(1∶5 000)和第二抗體山羊抗小鼠IgG(Goat Anti-Mouse lgG)(1∶20 000) 抗 體 進 行Western blot分析SeCDA7重組蛋白的表達。
1.2.3secda7基因的真核表達 設計帶有酶切位點的引物zSecda7 F和zSecda7 F進行secda7基因的克隆(表1),將secda7與pFastbac HTA真核表達載體進行連接,連接產物轉化JM109感受態細胞,挑取單菌落進行雙酶切鑒定。鑒定正確的重組質粒pFastbac HTA-secda7轉化DH10 BacTM感受態細胞,利用藍白板篩選方法篩選陽性克隆。鑒定正確重組Bacmid轉染昆蟲Sf9細胞,收集P3代病毒上清,分別利用第一抗體小鼠抗6×His單克隆抗體(Mouse anti-6×His tag monoclonal antibody)(1∶5 000)和第二抗體山羊抗小鼠IgG(Goat Anti-Mouse lgG)(1∶20 000)抗體進行Western blot分析SeCDA7重組蛋白的表達。
1.2.4secda7基因在甜菜夜蛾各組織中的表達分析 分別取甜菜夜蛾卵及預蛹,并將四齡甜菜夜蛾于冰上進行解剖,提取頭、表皮、中腸、圍食膜、馬氏管、脂肪體和蛻皮,利用RNeasy Mini Kit試劑盒(QIAGEN)提取總RNA,測定濃度后使用RevertAid First Strand cDNA Synthesis Kit試劑盒(Thermo Scientific)進行反轉錄合成cDNA。設計secda7基因相關熒光定量引物qSecda7 F和qSecda7 R(表1),以及以β-action為內參基因設計熒光定量引物(表1),參照Takara SYBR Premin Ex TaqTMⅡ說明書進行實時定量PCR。利用相對定量公式2-△△Ct進行數據分析。
1.2.5 重組蛋白SeCDA7幾丁質結合活性分析 通過Molano等[11]及Li等[12]的方法制備再生幾丁質,將10 g脫乙酰幾丁質粉末溶于350 mL 10%的乙酸,室溫過夜,次日,加入900 mL甲醇,利用玻璃纖維過濾后加入20 mL乙酸酐成凝膠狀,靜置30 min,用抹刀切片搗碎,用ddH2O洗至pH7.0。

表1 secda7基因全長引物及熒光定量引物
取1 mL純化后(利用NI-Agarose His標簽蛋白純化試劑盒純化)的重組SeCDA7蛋白與400 mg再生幾丁質在離心管中加入1 mmol/L PMSF室溫溫育,混勻后4℃過夜。次日,12 000 r/min離心3 min,棄上清,用1×PBS清洗2次,收集沉淀。將所得絡合物分別與PBS、2%SDS、2%SDS+5%β-巰基乙醇、0.5 mol/L氯化鈉、2%熒光增白劑和6 mol/L尿素室溫溫育15 min,離心后收集上清進行Western blot分析。
2 結果
2.1 secda7全長基因的獲得及序列分析
通過RT-PCR方法克隆得到編碼甜菜夜蛾幾丁質脫乙酰酶secda7基因。序列分析結果(圖1)顯示,secda7基因(GanBank登錄號為MG604929)全長1 431 bp,開放閱讀框為1 134 bp,編碼378個氨基酸,預測的SeCDA7蛋白的分子量和等電點約為43.156 kD和5.20。SignalP 4.1 Server進行信號肽分析顯示,在17和18氨基酸位點之間存在一個信號肽切割位點;NetNGlyc 1.0 Server軟件預測77和169氨基酸處存在兩個N-糖基化位點;結構域分析表明SeCDA7具有一個多聚糖乙酰基轉移酶催化區(氨基酸52-322位),屬于第Ⅴ類CDA蛋白。其氨基酸序列與鱗翅目昆蟲蓓帶夜蛾幾丁質脫乙酰酶(GenBank登錄號HM357864)具有較高的相似性。
2.2 重組SeCDA7的原核表達

圖1 secda7基因cDNA序列及推測的氨基酸序列
2.2.1 原核表達載體的構建secda7基因與pET-30a載體連接轉化BL21感受態細胞,成功構建重組菌株DH5α(pET-30a-secda7),經雙酶切鑒定,獲得5 422 bp載體和1 134 bp目的基因條帶,證明成功構建目的基因的原核表達載體,結果見圖2。原核表達載體的成功構建,可使攜帶外源核酸序列的重組載體進入大腸桿菌中進行目的蛋白的表達。

圖2 原核重組表達載體酶切鑒定結果
2.2.2 重組SeCDA7在大腸桿菌中的表達 用終濃度為1.25 mmol/L的IPTG在28℃對重組蛋白進行誘導,分別在誘導2、4、6、8 h取樣,收集菌體沉淀進行檢測。Western blot 分析重組SeCDA7蛋白在大腸桿菌中的表達,結果見圖3,在28℃,用1.25 mmol/L IPTG誘導2 h SeCDA7蛋白表達量最高,蛋白大小約為47 kD。
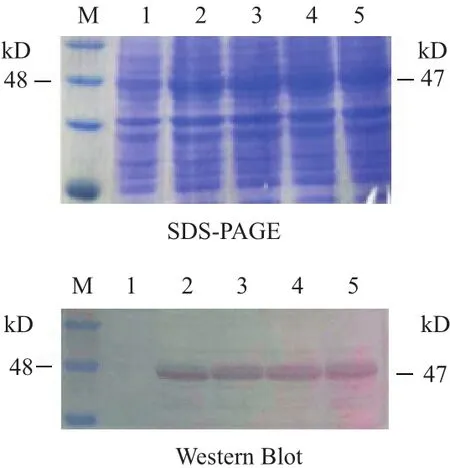
圖3 SeCDA7的SDS-PAGE及Western Blot表達圖譜
2.3 重組SeCDA7的真核表達
2.3.1 真核表達載體的構建 將secda7基因與pFastbac HTA載體連接轉化JM109感受態細胞,成功構建重組菌株JM109(pFastbac HTA-secda7),瓊脂糖凝膠電泳(圖4)顯示,出現4 776 bp載體大小條帶和1 134 bp目的基因大小條帶,證明成功構建目的基因的真核表達載體。將重組pFastbac HTA-secda7質粒轉入E.coliDH10BacTM感受態細胞,藍白斑篩選得到重組Bacmid質粒,經PCR鑒定,得到3 564 bp大小的條帶,表明成功得到重組Bacmid質粒。
2.3.2 重組SeCDA7在Sf9昆蟲細胞的表達 將重組Bacmid轉染生長狀態良好的Sf9昆蟲細胞,昆蟲細胞出現明顯感染癥狀,細胞直徑和細胞核增大,視野中出現顆粒體(圖5),得到P3代病毒培養72 h時,Western blot分析(圖6)顯示SeCDA7在昆蟲細胞中表達大小約為47 kD的目的蛋白。
2.4 secda7基因在甜菜夜蛾各組織中的表達分析
實時定量PCR結果(圖7)顯示,secda7基因在甜菜夜蛾的卵、預蛹、頭、表皮、中腸、馬氏管及脂肪體中有表達,但是表達量較低,在中腸中表達量最高,為中腸特異性表達蛋白。

圖4 重組桿狀病毒轉座載體的鑒定
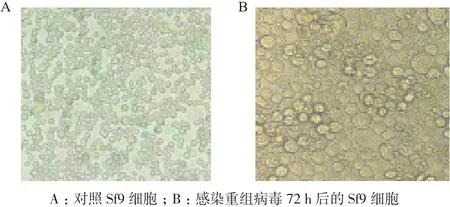
圖5 重組病毒轉染前后細胞狀態比較(10×)

圖6 SeCDA7表達產物的Western blot分析
2.5 SeCDA7幾丁質結合活性測定
外源表達的SeCDA7蛋白表現出幾丁質結合活性,與鱗翅目昆蟲粉紋夜蛾Tn-P42蛋白類似,在體外與幾丁質可緊密結合,Western blot分析結果(圖8)顯示,SeCDA7蛋白僅用強洗脫劑2%SDS+5%β-巰基乙醇和6 mol/L尿素以及2%熒光增白劑可將其洗脫,而用PBS、2%SDS、0.5 mol/L氯化鈉不能將其洗脫。

圖7 secda7基因在甜菜夜蛾不同組織中的表達模式

圖8 SeCDA7重組蛋白幾丁質結合活性Western blot分析
3 討論
昆蟲幾丁質脫乙酰酶(Chitin deacetylase,CDA)是昆蟲體內一類重要幾丁質結合蛋白,可分布在蟲體內多個含幾丁質成分組織結構中,是幾丁質降解酶系成員之一,能夠催化幾丁質中β-1,4糖苷鍵連接的N-乙酰基葡糖胺的乙酰胺基水解,將幾丁質修飾為殼聚糖,在昆蟲生長、發育和代謝中具有重要作用[13-15]。從2005年以來,研究者從不同的昆蟲中開始探索CDA類蛋白的生物功能,這對昆蟲生物防治具有重要生物學意義。
SeCDA7蛋白成功在大腸桿菌中表達,有利于研究蛋白純化、定位及功能分析。利用pFastBac HT A載體使得SeCDA7蛋白成功在Sf9昆蟲細胞中表達,并對其進行純化與表達分析,并鑒定出SeCDA7蛋白具有幾丁質結合活性,這與粉紋夜蛾(Trichoplusia ni)[6]、華北大黑鰓金龜(Holotrichia oblitaFaldermann)[16]第Ⅴ類CDA蛋白表現的幾丁質結合活性相一致。通過熒光定量PCR技術,對secda7基因進行了轉錄水平的組織定位,屬于中腸特異蛋白,這與第Ⅴ類CDA蛋白功能具有直接相關性。
SeCDA7蛋白屬于昆蟲中腸特異蛋白,圍食膜作為昆蟲體內抵御外源性物質的第一道天然屏障,具有保護中腸上皮細胞,阻止病原物的侵染等多種功能,圍食膜主要由蛋白質、多糖和幾丁質構成,其中蛋白質對于維持圍食膜的致密結構至關重要,推測第Ⅴ類CDA蛋白與圍食膜結構功能緊密相關,因此圍食膜蛋白是進行害蟲防治重要的切入點之一。對幾丁質脫乙酰酶功能的研究不只局限于昆蟲中,在一些真菌、細菌及蟹類、魚類等海洋生物都有相關研究報道。在尼杜拉曲霉、苜蓿中華根瘤菌、構巢曲霉和稻瘟病菌中也分離得到不同幾丁質脫乙酰酶類[17-20],參與不同的生理功能;在線蟲中,也得到了幾丁質脫乙酰酶類蛋白[21],在對藥材甲蟲的研究中,表明SpCDA1對于S. paniceum中幼蟲到蛹轉變是必不可少的[22],在對紅雪蟹的研究中,僅在表皮中發現CDA存在,推測可能與蛻皮相關[23]。幾丁質脫乙酰酶類蛋白的廣泛研究,明確幾丁質脫乙酰酶類蛋白在昆蟲生長發育中的作用及深入探究幾丁質脫乙酰酶的生理功能也顯得尤為重要。
4 結論
克隆了甜菜夜蛾幾丁質脫乙酰酶secda7全長基因,在原核和真核中表達約47 kD的蛋白,生物信息學分析結果顯示SeCDA7含有一個結構域,屬于第Ⅴ類CDA蛋白,Western Blot分析結果顯示SeCDA7蛋白具有幾丁質結合活性,熒光定量PCR結果表明secda7基因屬于中腸特異蛋白。
