磷濃度對海水鈍頂螺旋藻生長代謝的影響
2019-08-08 03:32:02陳浩楊冰潔李濤吳華蓮吳后波向文洲
生物技術通報 2019年8期
關鍵詞:影響
陳浩 楊冰潔 李濤 吳華蓮 吳后波 向文洲
(1. 中國科學院南海海洋研究所 中國科學院熱帶海洋生物資源與生態重點實驗室,廣州 510301;2. 中國科學院大學,北京 100049)
螺旋藻(Spirulina)或節旋藻(Arthrospira)為藍藻門(Cyanophyta)、段殖藻目(Hormogonales)、顫藻科(Oscillatoriaceae)、螺旋藻屬(Spirulina)或節旋藻屬(Arthrospira)的藻類。螺旋藻是目前蛋白質含量最高的食品資源[1],同時,螺旋藻還富含功能獨特的活性多糖、藻藍蛋白、γ-亞麻酸等生物活性物質,在食品、醫藥保健、化妝品及飼料等領域得以廣泛應用[2]。研究表明,螺旋藻多糖對HIV病毒、丙肝病毒等多種病毒具有抑制作用[3],而且其在抗腫瘤、抗氧化[4]、免疫調節、降血糖血脂以及修復DNA損傷等方面也表現出了良好的功效[5-6]。螺旋藻還含有豐富的藻藍蛋白,藻藍蛋白已被證實具有顯著的抗癌、抗炎癥等多種生物活性[7],同時還能作為食用色素應用于食品行業[8]。γ-亞麻酸對心血管疾病具有良好預防作用,而且是合成前列腺素E1的前體物質[9]。
螺旋藻產業在國際上興起于20世紀80年代初,我國于90年代中期已成為國際上最大的螺旋藻產業國[10]。利用天然海水培養螺旋藻,可顯著節約淡水資源,同時可利用灘涂、鹽堿地等環境進行培養,有效緩解農用耕地日趨緊張的問題,而海水中含有大量營養元素,可大幅度降低螺旋藻培養的肥料成本。長期以來,海水螺旋藻的研究得到全世界的普遍關注,但目前大多僅停留在實驗室藻種選育階段。本實驗室于20世紀80年代末至90年代初在國際上率先建立了全海水的螺旋藻產業化生產技術,并與企業合作成功實現了海水螺旋藻的產業化,與傳統淡水螺旋藻養殖相比,其品質得以顯著提高[11-12]。
磷是調節細胞生長和代謝的必需營養因子之一,在大多數細胞過程中起著不可或缺的作用,特別是涉及能量轉移、信號轉導、高分子生物合成、光合作用和呼吸的過程[13]。研究表明磷元素對許多藻類脂質[14-15]、蛋白質[16]和多糖[17-18]的積累具有顯著影響。螺旋藻的主要成分及其含量可以通過多種培養條件來控制,如氮濃度[19]、光照強度[20]和溫度[21]。有關氮源、溫度和光照對螺旋藻的生長、代謝產物影響研究十分廣泛,但磷源濃度對螺旋藻生長代謝影響的研究還很少,僅見2篇文獻對淡水螺旋藻的主要生化組分[22]和脂類積累[23]影響的研究。對于磷濃度對海水螺旋藻生物量生產和生化組成的影響,尚未見報道。因此,研究磷對海水螺旋藻生長和生化組成的影響對海水螺旋藻主要高值化產物生產以及海水螺旋藻產業具有重要的指導意義。本研究首次開展不同磷源濃度的海水螺旋藻培養實驗,系統分析磷源濃度對海水螺旋藻生長及生化組成的影響規律,為通過磷調節來提高螺旋藻主要產物的產量、提升螺旋藻品質奠定理論基礎。
1 材料與方法
1.1 材料
1.1.1 藻株及保存條件 海水鈍頂螺旋藻(Spirulina Platensis)藻種,由中國科學院南海海洋研究所海藻資源與生物技術實驗室提供。
藻種保存條件:培養溫度為25±1℃,熒光燈提供光源,培養光強為 49-57 μmol·photons m-2·s-1,光暗周期為 12 h∶12 h。
1.1.2 實驗設計 本實驗以改良的Zarrouk海水培養基為基礎培養基,其組成為:5 g/L NaHCO3,0.5 g/L NaNO3,0.01 g/L FeSO4·7H2O,0.08 g/L Na2EDTA,1 mL/L A5solution,以天然海水為基質配制而成。通過補加去離子水將鹽度控制在28±1‰。K2HPO4·3H2O為磷源,加入不同的磷源濃度(終濃度分別為 0.005、0.01、0.02,0.03和 0.04 g/L),接種于500 mL錐形瓶,于溫度為25℃,光強為49-57 μmol·photons m-2·s-1條件下,靜置培養,每天定時搖瓶6次。培養結束后(培養周期10 d),離心收集藻泥,去離子水多次清洗,真空冷凍干燥獲得凍干粉,用于總糖、水溶性多糖、蛋白質、藻藍蛋白、光合色素總脂以及脂肪酸含量的測定。
1.2 方法
1.2.1 生物質濃度的測定 利用干重法進行測定。取一定量的藻液,用預先在80℃烘箱中烘干至恒重的混合纖維濾膜(0.45 μm)進行抽濾,再將有藻細胞的濾膜放置在80℃烘箱烘至恒重,用減差法得到微藻細胞干重。每個樣品重復3次計算平均值。
1.2.2 總糖的提取及測定 取10 mg 藻粉,加入5 mL 0.5 mol/L H2SO4,于80℃ 水浴攪拌0.5 h,8 000 r/min 離心10 min,收集上清液。反復抽提3次,合并上清后定容到50 mL,得到總糖提取液;總糖含量的測定采用苯酚-硫酸法[24]。
1.2.3 水溶性多糖提取及測定 取50 mg 藻粉溶于水,超聲破壞細胞結構后,70℃熱水浸提4 h,8 000 r/min 離心10 min 收集上清。反復抽提4次,合并上清并定容至50 mL得到水溶性多糖提取液;水溶糖含量通過苯酚-硫酸法測定。
1.2.4 胞壁多糖及總多糖的測定 取上述提取水溶性多糖后所得藻渣,加入5 mL 0.5 mol/L H2SO4,于80℃ 水浴攪拌 0.5 h,8 000 r/min 離心 10 min,收集上清液。反復抽提3 次,合并上清后定容到50 mL,得到胞壁多糖提取液[25];采用苯酚-硫酸法測定其含量。
1.2.5 蛋白質含量的測定 采用半自動凱氏定氮儀測定總蛋白質含量,以蛋白質的F值為6.25 計算。
1.2.6 藻膽蛋白的測定 采用改良的Sigelman和Kycia 法提取藻膽蛋白,分光光度法測定藻藍蛋白、異藻藍蛋白含量[26]。
1.2.7 脂溶性光合色素測定 取10 mg 藻粉,置于10 mL玻璃離心管中,加入5 mL丙酮,避光冰浴攪拌提取1-2 d,直至藻體變白,3 000 r/min 離心10 min 收集上清得到色素提取液。采用分光光度法在662 nm、645 nm和470 nm下測定葉綠素a、總類胡蘿卜素含量[27-28],由下列公式計算:

色素含量(%DW)=(色素質量濃度算×提取液體積)/藻體干重
1.2.8 總脂的測定與分級 總脂的提取與含量的測定采取改良的Khozin-Goldberg方法[29]。
1.2.9 脂肪酸組成的測定 稱取25 mg凍干藻粉于10 mL玻璃離心管中,加入2 mL 2% H2SO4無水甲醇:甲苯(9∶1,V∶V),充入氮氣后,用80℃水溶攪拌1 h,再依次加入1 mL去離子水和1 mL正己烷,充分震蕩后3 500 r/min離心5 min,將上層有機相轉移至另一離心管中,氮氣吹干,再加入1 mL含有C17標準品的正己烷,并用孔徑為0.22 μm的濾膜過濾至1.5 mL的小玻璃瓶中,最后利用氣相色譜儀測定脂肪酸。
1.2.10 統計分析 數據以平均值,最后利用氣相色譜SPSS13.0 統計軟件進行方差分析,檢驗水平儀=0.05,P<0.05差異有統計學意義。使用Origin 8.1進行數據處理和圖表制作。
2 結果
2.1 磷源濃度對海水鈍頂螺旋藻生長的影響
經過0-2 d的生長延滯期,從第2天開始,海水鈍頂螺旋藻在5種不同磷源濃度中均開始迅速增長,至第10天培養結束時,0.005 g/L、0.01 g/L、0.02 g/L、0.03 g/L和0.04 g/L 5種磷源濃度處理組的生物質濃度分別達到0.412±0.003 g/L、0.416±0.003 g/L、0.433±0.002 g/L、0.450±0.016 g/L 和 0.484±0.022 g/L,如圖1所示。結果顯示,隨著磷源濃度的增加,海水螺旋藻的生物質濃度也隨之提高,濃度為0.04 g/L處理組的生物質濃度高于其他磷源濃度處理組,為0.484±0.022 g/L。由方差分析可知,除0.005 g/L處理組與0.01 g/L處理組無顯著差異外(P>0.05),其余處理組生物質濃度之間均存在顯著差異(P<0.01)。

圖1 磷源濃度對海水鈍頂螺旋藻生長的影響
2.2 磷源濃度對海水鈍頂螺旋藻蛋白質含量的影響
對第10天不同磷源濃度影響下海水鈍頂螺旋藻蛋白質含量進行測定。如圖2所示,經方差分析后得出在磷源濃度為0.02 g/L和0.03 g/L時,海水螺旋藻蛋白質含量無顯著差異(P>0.05),其余處理組均存在顯著差異(P<0.01)。由此可見,隨著磷源濃度的增加,海水鈍頂螺旋藻蛋白質含量顯著升高,最高可達59.23±0.61%DW。海水鈍頂螺旋藻蛋白質產量變化與含量變化一致,即隨著磷源濃度增加而升高,并在磷源濃度為0.04 g/L時,達到最高值0.29±0.02 g/L。

圖2 磷源濃度對海水鈍頂螺旋藻蛋白質產量的影響
2.3 磷源濃度對海水鈍頂螺旋藻多糖含量的影響
對第10天不同磷源濃度影響下海水鈍頂螺旋藻的多糖組成及含量測定。如圖3所示,經方差分析可得在磷源濃度為0.02 g/L、0.03 g/L和0.04 g/L時,海水鈍頂螺旋藻總糖含量均無顯著差異(P>0.05),其余各處理組均有顯著差異。由此可見,隨著磷源濃度的增加,海水螺旋藻總糖含量顯著下降后達到平穩,濃度0.005 g/L處理組總糖含量達到最高(25.96±1.61%DW)。其總多糖產量變化與含量變化相似,濃度0.005 g/L處理組含量最高(107.05±2.83 mg/L),如下圖3所示。
如表1所示,海水螺旋藻多糖主要為水溶性多糖,磷源濃度0.02 g/L、0.03 g/L和0.04 g/L組,海水鈍頂螺旋藻水溶糖以及胞壁糖含量均無顯著差異(P>0.05),其余各處理組均有顯著差異(P<0.01)。兩個組分的含量隨著磷源濃度的升高而降低,磷源濃度0.005 g/L處理組含量最高,分別為22.15±0.71%DW以及3.62±0.28%DW。磷源濃度0.02-0.04 g/L范圍內,隨著磷源濃度增加,兩個多糖組分的含量趨于穩定,不再隨著濃度增加而顯著變化(P>0.05)。

圖3 磷源濃度對海水鈍頂螺旋藻多糖產量的影響
2.4 磷源濃度對海水鈍頂螺旋藻光合色素的影響
對第10天不同磷源濃度條件下海水螺旋藻的藻藍蛋白含量進行測定。如圖4所示,經方差分析可知,磷源濃度0.02 g/L和0.03 g/L處理組的海水螺旋藻C-藻藍蛋白、別藻藍蛋白、總藻膽蛋白含量無顯著差異(P>0.05);磷源濃度0.01 g/L和0.04 g/L處理組,海水螺旋藻C-藻藍蛋白、別藻藍蛋白和總藻膽蛋白含量無顯著差異(P>0.05),其余各處理組均有顯著差異(P<0.01)。由此可見,隨著磷源濃度的增加,C-藻藍蛋白、別藻藍蛋白和總藻膽蛋白含量升高,但隨著磷源濃度進一步增加,3個組分含量略有下降。磷源濃度0.02 g/L處理組達到最大值,3種組成含量分別為14.56±0.99%DW、4.21±0.19%DW和18.77±0.39% DW。
如圖5所示,藻膽蛋白各組分產量隨著磷源濃度增加呈增長趨勢,磷源濃度0.04 g/L處理組的C-藻藍蛋白、別藻藍蛋白及總藻膽蛋白產量最高,分別達到0.064±0.001 g/L、0.019±0.001 g/L及0.084±0.001 g/L,但與0.02 g/L和0.03 g/L處理組無顯著差異(P>0.05)。
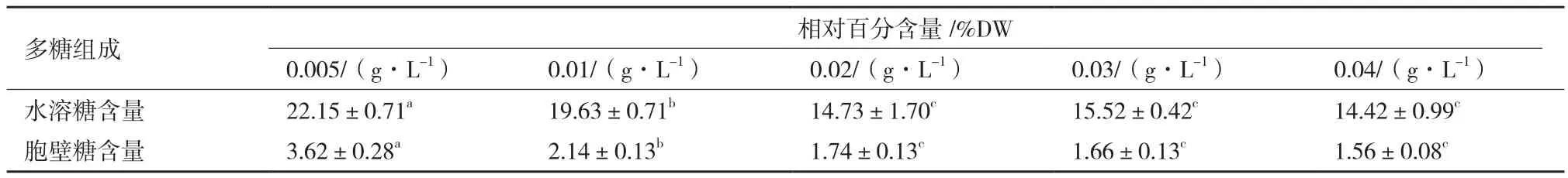
表1 磷源濃度對海水鈍頂螺旋藻多糖含量及組成的影響

圖4 磷源濃度對海水鈍頂螺旋藻藻膽蛋白組成及含量的影響

圖5 磷源濃度對海水鈍頂螺旋藻藻膽蛋白組成及產量的影響
如表2所示,不同處理組海水螺旋藻葉綠素a含量均存在顯著差異(P<0.01);磷源濃度0.02 g/L、0.03 g/L、0.04 g/L處理組的類胡蘿卜素含量沒有顯著差異(P>0.05),其余各處理組均存在顯著差異(P<0.01)。海水螺旋藻葉綠素a含量隨著磷源濃度的增加由0.46±0.005%上升至1.01±0.010%,磷源濃度0.04 g/L處理組含量最高。螺旋藻類胡蘿卜素含量隨磷源濃度增加有所升高,但變化幅度不大,磷源濃度達0.02 g/L時趨于平穩,類胡蘿卜素含量為0.35±0.018%。總體而言,螺旋藻葉綠素a含量隨磷源濃度增加而升高,磷源濃度0.04 g/L處理組含量最高,類胡蘿卜素含量隨磷源濃度增加先升高后平穩。
2.5 磷源濃度對海水鈍頂螺旋藻總脂的影響
對第10天不同磷源濃度條件下海水螺旋藻的總脂含量進行測定。如圖6所示,磷源濃度0.005 g/L、0.01、0.02、0.03及0.04 g/L處理組的總脂含量分別為7.22±0.68%DW、6.89±0.50% DW、7.94±0.88%DW、7.38±1.02% DW以及8.49±0.76% DW經方差分析可知,在磷源濃度的影響下,各處理組海水鈍頂螺旋藻總脂含量差異并不顯著(P>0.05)。因此,海水鈍頂螺旋藻總脂含量受磷源濃度影響不大。

圖6 磷源濃度對海水鈍頂螺旋藻總脂含量的影響

表2 磷源濃度對海水鈍頂螺旋藻光合色素含量的影響
2.6 磷源濃度對海水鈍頂螺旋藻脂肪酸組成的影響
對第10天不同磷源濃度條件下海水螺旋藻的脂肪酸組成進行測定。如表3所示,海水鈍頂螺旋藻脂肪酸組分結構較為簡單,碳鏈長度多為C16-C18的脂肪酸,其飽和脂肪酸主要為C16∶0,多不飽和脂肪酸主要為 γ-亞麻酸(C18∶3,ω-6),其 γ-亞麻酸相對百分含量并無明顯規律,磷源濃度0.005與0.03 g/L處理組無顯著差異(P>0.05),磷源濃度0.01、0.02與0.04 g/L處理組也不存在顯著性差異(P>0.05),但0.005與0.03 g/L處理組 γ-亞麻酸相對百分含量顯著高于其他處理組,但最大相對差異僅3.7%,升高幅度不大。

表3 磷源濃度對海水鈍頂螺旋藻脂肪酸相對百分含量的影響
3 討論
3.1 磷源濃度對海水鈍頂螺旋藻生長及產物積累的影響
由于在產業應用中,螺旋藻多采取半連續培養,采收時間選擇指數生長期末期而非生長減緩的平臺期,可獲得更高的生物量產率[30],因此,為了更切合生產應用,本研究對處于指數生長末期第10天的海水鈍頂螺旋藻進行采收和生化指標測定。
本研究表明,磷源濃度對海水鈍頂螺旋藻的生長及生化組分有著重要影響。海水鈍頂螺旋藻的生長隨著磷源濃度的增加而升高,在磷源濃度為0.04 g/L時,達到最高值。這一點海水鈍頂螺旋藻與一些微藻類似,如底棲硅藻[31]等。但也有一些微藻在過高的磷源濃度下生長反而受到抑制,如柵藻在磷源濃度0.2 mg/L時藻細胞濃度達到最大,磷源濃度的進一步增加使柵藻生長受到抑制[32]。由此可以看出海水鈍頂螺旋藻能夠耐受較高范圍的磷源濃度。
低磷條件下海水鈍頂螺旋藻優先合成多糖,這一結果與Markou等[16]在淡水螺旋藻中的結果相符。隨著磷源濃度的增加,蛋白質、藻藍蛋白、葉綠素和類胡蘿卜素的產量都有顯著的升高。因此在生產應用中,可以通過選擇較高的磷源濃度促進這些產物的合成積累。如需獲得多糖含量高的螺旋藻粉則可以通過降低磷源濃度來實現。
本研究探究了磷對螺旋藻藻膽蛋白含量的影響,結果表明,隨著磷源濃度的增加,海水鈍頂螺旋藻的C-藻藍蛋白和別藻藍蛋白的含量均出現先增加后下降的趨勢,目前尚未有開展螺旋藻相關研究的其他報道。同時,由于隨著磷源濃度的增加而增加,藻藍蛋白的產量增加,但磷源濃度達到0.02 g/L后,3組高濃度處理的差異不顯著(P>0.05),因此,0.02 g/L的磷源可能是比較適宜藻藍蛋白積累的合理濃度。
3.2 磷元素對螺旋藻生長代謝影響的生理機制探討
磷源是微藻生長的必須元素之一,在細胞代謝中,磷是ATP、RNA、DNA、生物膜磷脂等重要生化組分的重要元素[33],與細胞生長、分裂、遺傳變異、能量代謝、信號傳遞和抗逆性等關鍵生命過程有關。磷是多種酶的輔基,不僅參與碳水化合物的代謝以及脂肪的轉化,而且也是葉綠素合成所必需的[34]。由圖2和圖3中我們可以看出,在海水螺旋藻中多糖含量與蛋白質含量有著類似互補的關系,在低磷條件下,多糖含量最高,蛋白質含量最低,高磷條件則相反。
控制碳水化合物合成的關鍵酶ADP葡萄糖焦磷酸化酶可被高濃度無機磷抑制,因而無機磷濃度增加會導致碳水化合物合成受阻[35-36]。這一機制可能是本研究中海水螺旋藻多糖含量隨磷濃度增加而顯著降低的原因之一。
同時,低磷條件下導致RNA,甚至包括DNA和ATP的含量急劇下降[37],蛋白質含量也隨之減少,應為低磷條件下多糖含量高的另一個原因。Dean等[38]與 Lynn等[39]報道磷饑餓會導致Anabaena flos-aquae與Stephanodiscus minutulus生物量組成碳水化合物積累,抑制蛋白質合成,與本研究結果相符。同樣,磷對光合色素含量的影響可能與磷對蛋白質(酶)的變化相關,因此總體上呈現類似的變化規律。此外,光合色素的變化反過來也影響蛋白質、多糖和能量代謝過程,也不排除其他生理過程的參與。
綜上所述,本研究表明,磷對海水鈍頂螺旋藻重要產物積累影響十分顯著,通過磷源調節可有效定向誘導海水螺旋藻主要高值化產物的合成積累。同時,磷對海水鈍頂螺旋藻生長代謝的影響規律也較為復雜,其生理代謝機制有待更加深入地探索,對更好地發展螺旋藻產業極為重要。
4 結論
低濃度的磷源可促進海水螺旋藻多糖合成,磷源濃度增加有利于提高海水螺旋藻蛋白質、藻藍蛋白、光合色素含量。通過磷源可誘導海水螺旋藻不同的高值化產物的積累,對海水螺旋藻產業化培養與開發具有指導意義。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
