結核分枝桿菌KatG-ACT271CCT(Thr271Pro)突變與異煙肼耐藥關系的研究
2019-07-16 08:21:58楊壽慧1劉惟優吳龍火3袁小亮
中國人獸共患病學報 2019年6期
關鍵詞:耐藥
楊壽慧1,劉惟優,吳龍火3,袁小亮
結核病(Tuberculosis,TB)長期以來威脅著人類的健康,世界衛生組估計2015年全世界約有1 040萬新發結核病例,且約有48萬新發耐多藥結核(multidrug-resistant,MDR)病例,我國新發肺結核人數僅次于印度和印度尼西亞而位居全球第3位[1]。異煙肼(isoniazid,INH)是結核病治療的首選化學藥物。然而,隨著INH廣泛用于臨床,其耐藥情況不容忽視,據2010年全國第5次結核病流行病學抽樣調查報告[2],我國結核病患者對INH的耐藥率最高(28.6%)。這一嚴峻的形勢迫使人們尋求更加快速、準確的INH耐藥結核檢測方法,以便更有效的控制結核病。
目前認為,katG基因(編碼過氧化氫-過氧化物酶)是最重要的INH耐藥基因,其突變存在于50%~70%的耐INH結核菌株中[3]。故katG基因突變檢測已成為早期判斷耐INH結核菌的重要手段。而katGAGC315ACC(Ser315Thr)突變是katG基因最常見的突變類型[4]。課題組前期研究發現來自江西地區的一臨床分離結核菌株對INH表現高度耐藥,經測序發現此菌株在katG基因含有唯一突變點ACT271CCT(GenBank登錄號:JN049496),但該突變點與INH耐藥關系尚無相關研究報道[5]。本研究采用katG蛋白酶活性檢測及分子對接方法分析該突變點與INH耐藥的關系,以期通過檢測發現該位點基因突變與結核菌株表型耐藥關系。
1 材料與方法
1.1菌株和質粒 結核分枝桿菌katG(ACT271CCT)、katG野生型、katG(AGC315ACC)基因片段由上海生工生物工程合成,BL21(DE3)pLySs贛南醫學院基礎醫學院科研中心保存。質粒pET-22b(+)購自武漢淼靈生物科技有限公司。
1.2主要試劑 酵母粉、胰蛋白胨購自英國OXOID公司,HisTrap FF-crude購自GE公司,氨芐青霉素、Anti-His antibody、辣根過氧化物酶標記山羊抗鼠IgG、過氧化氫酶(catalase, CAT)檢測試劑盒及過氧化物酶試劑盒(peroxidase, POD)購自索萊寶科技有限公司。PageRulerTMPrestained Protein ladder購自Thermo Fisher公司。
1.3 結核分枝桿菌katG表達及提純
1.3.1原核表達載體的構建 katG野生型、katG(ACT271CCT)、katG(AGC315ACC)基因片段和載體pET-22b(+)分別用EcoRⅠ和HindⅢ進行雙酶切。酶切產物經DNA連接酶與載體pET-22b(+)連接。
1.3.2katG重組蛋白誘導表達條件摸索 將各重組質粒及pET-22b(+)轉化入BL21(DE3)pLySs感受態,接種于含100 μg/mL氨芐青霉素的SOB平板,經37 ℃過夜培養。無菌條件下挑取轉化陽性單克隆菌接種于SOB培養液中,待A600值約為0.6 時,按1∶100轉接至新的SOB培養液中。取野生型培養重組質粒菌液培養3~5 h后,摸索不同IPTG濃度、溫度、時間、培養液濃度下蛋白誘導表達最佳條件。
1.3.3katG重組蛋白原核表達及Western blot鑒定 按蛋白誘導表達最佳條件,分別誘導katG野生型、katG(ACT271CCT)、katG(AGC315ACC)重組質粒菌,次日蛋白表達充分后,分別取等量菌液離心,用電泳液重懸菌體,行SDS-PAGE電泳及Western blot鑒定。Western blot一抗為Anti-His antibody,二抗為辣根過氧化物酶標記山羊抗鼠IgG,采用辣根過氧化物酶法(HRP)顯色。
1.3.4katG重組蛋白分離、純化 分別重新從SOB平板上挑取各自單克隆菌,接種于10 mL SOB培養液中,過夜培養后,轉接至新的500 mL SOB培養液中,過夜誘導蛋白表達。收集菌體,用20 mL 20 mmol/L PBS+10 mmol/L咪唑平衡緩沖液重懸。菌液加入溶菌酶(工作濃度0.2~0.4 mg/mL),超聲破碎儀破菌,10 000 r/min 4 ℃ 20~30 min分離上清和沉淀,分別取上清和沉淀進行SDS-PAGE電泳,分析超聲破碎后katG重組蛋白分布情況。
將HisTrap FF-crude柱用20 mmol/LBS沖洗酒精后,用20 mmol/L PBS+10 mmol/L咪唑平衡緩沖液平衡5個柱長體積,將超破后上清以1 mL/min通過柱子,過柱2遍,使目的蛋白與柱子充分結合,收集流出液,分別用含20、40、100、250 mmol/L咪唑緩沖液依次洗脫His標簽純化樹脂,收集各組洗脫液進行SDS-PAGE電泳,分析katG重組蛋白純化情況。將重組蛋白液加入透析袋中,置于透析液中4 ℃過夜透析。
1.3.5katG重組蛋白定量及活性測定 用BCA蛋白定量法檢測katG重組純化蛋白濃度。按過氧化氫酶試劑盒說明書比例混合好工作液,在1 000 μL過氧化氫酶反應體系中加入100 μL重組蛋白,紫外分光光度計240 nm測定初使吸光值A1和1 min后的吸光值A2。ΔA=A1-A2,計算katG重組蛋白過氧化氫酶活性(U/mg prot)=678×ΔA÷蛋白濃度。按過氧化物酶試劑盒說明書比例混合好工作液,在1 000 μL過氧化物酶反應體系中加入100 μL重組蛋白,分光光度劑470 nm檢測加入重組蛋白30 s 時的吸光值 A1和 90s 時吸光值 A2, ΔA=A2-A1,計算katG重組蛋白過氧化物酶活性(U/mg prot)=7 133×ΔA÷蛋白濃度。
1.4分子對接 野生型結核分枝桿菌katG蛋白的晶體結構從蛋白晶體結構數據庫 (http://www.rcsb.org)獲得(PDB ID:1SJ2),使用Autodock Tools對受體蛋白質結構進行處理,保留其氧化反應中心含鐵的卟啉環,刪除底物小分子,加極性氫,加原子電荷。其他突變體均在野生型的基礎上通過YASARA軟件進行突變然后使用最陡下降法優化1 000次選擇自由能最小的構象建立katG蛋白三維模型。
異煙肼的三維結構從PubChem compound數據庫(ID:3767)獲取,將上述的小分子結構使用分子力場方法優化,并加上Gaisteiger電荷,作為對接配體存為pdbqt文件。然后對小分子和受體進行分子對接,所有對接均使用Autodock 4.2軟件包完成,利用拉馬克遺傳算法(Lamarckian Genetic Algorithm)確定katG 蛋白和異煙肼最佳結合構象,對接結果保留10個能量最低的構象并進行簇(cluster)分析。參考氧化活性中心卟啉環的位置,對接的 BOX位于卟啉上方的空腔內,中心坐標為45.47,-3.905, 28.640,大小為 40埃×40埃×40埃,其余采用軟件的默認值。
2 結 果
2.1 katG蛋白表達、純化及酶活性檢測
2.1.1katG蛋白表達條件摸索 結核分枝桿菌katG野生型重組基因轉化至BL21(DE3)pLySs,摸索不同溫度、不同濃度IPTG、不同濃度培養基,IPTG誘導katG蛋白表達最佳條件。可以看出IPTG 0.5 mmol/L、20 ℃、12 h在2×SOB培養基條件下誘導出目的蛋白濃度最佳(圖1、2)。
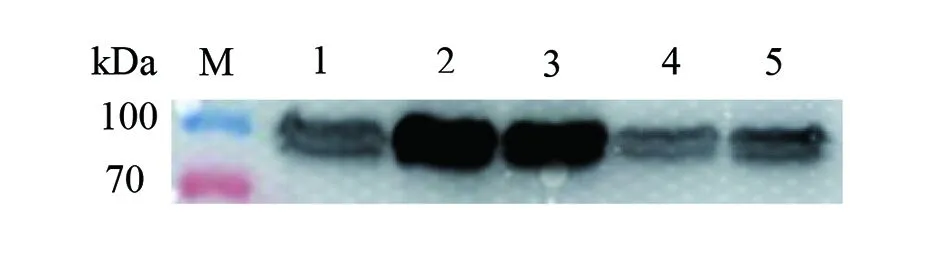
M:蛋白分子質量標準;1:未加IPTG、28 ℃、12 h;2:IPTG 0.5 mmol/L、20 ℃、12 h;3:IPTG 0.5 mmol/L、25 ℃、12 h;4:IPTG 0.5 mmol/L、28 ℃、12 h;5:IPTG 1 mmol/L、28 ℃、12 h。圖1 IPTG濃度、時間、溫度的摸索Fig.1 Exploration of IPTG concentration,induction time and temperature
M:蛋白分子質量標準;1:未轉質粒大腸桿菌;2:轉質粒菌體誘導于1×SOB培養基;3:轉質粒菌體誘導于1.5×SOB培養基;4:轉質粒菌體誘導于2×SOB培養基。圖2 培養基濃度的摸索Fig.2 Exploration of medium concentration
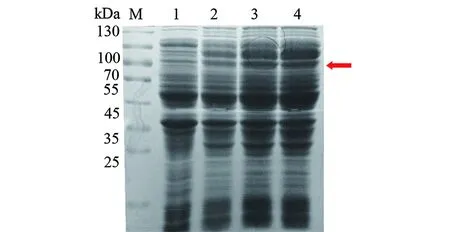
2.1.2katG重組蛋白原核表達及Western blot鑒定 結核分枝桿菌katG野生型、katG(ACT271CCT)、katG(AGC315ACC)重組基因轉化至BL21(DE3)pLySs,IPTG誘導重組基因蛋白表達,重組蛋白分子量約為85 kd,SDS-PAGE檢測誘導前后蛋白表達情況(圖3)。Western blot顯示含有6×His標簽的重組蛋白能被Anti-His antibody識別(圖4)。
2.1.3超聲破碎重組蛋白大腸桿菌 重組基因菌體超聲破碎后katG蛋白分布情況可見katG蛋白分布于上清中(圖5)。
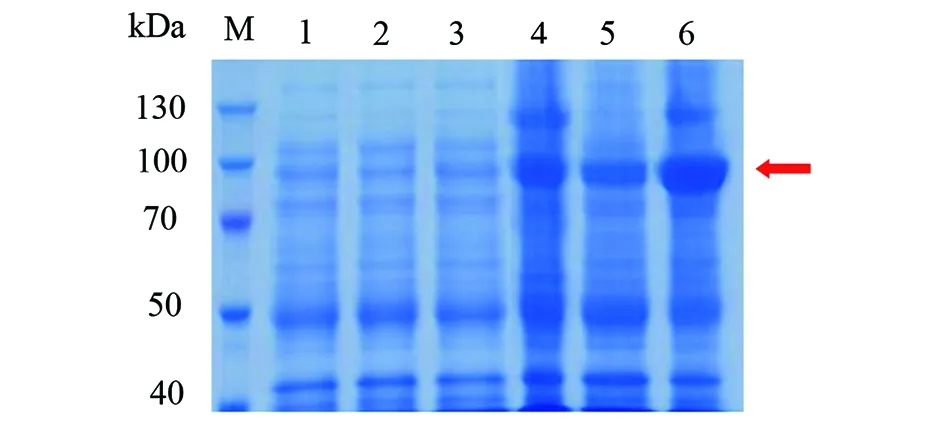
2.1.4katG重組蛋白的純化 重組菌超聲破碎上清液過HisTrap FF-crude柱,不同濃度咪唑洗脫緩沖液洗脫后檢測,其中100 mmol/L咪唑洗脫緩沖液得到純化katG重組蛋白(圖6)。
2.1.5katG重組蛋白酶活性測定 測得katG(ACT271CCT)、katG野生型、katG(AGC315ACC)重組蛋白過氧化氫酶和過氧化物酶活性,見表1和圖7。
M:蛋白分子質量標準;1:katG蛋白野生型未誘導;2:katG蛋白271突變未誘導;3:katG蛋白315突變未誘導;4:katG蛋白野生型誘導后;5:katG蛋白271突變誘導后;6:katG蛋白315突變誘導后。圖3 重組表達宿主菌誘導前后重組蛋白katG的表達Fig.3 Recombinant protein katG expression before and after induction
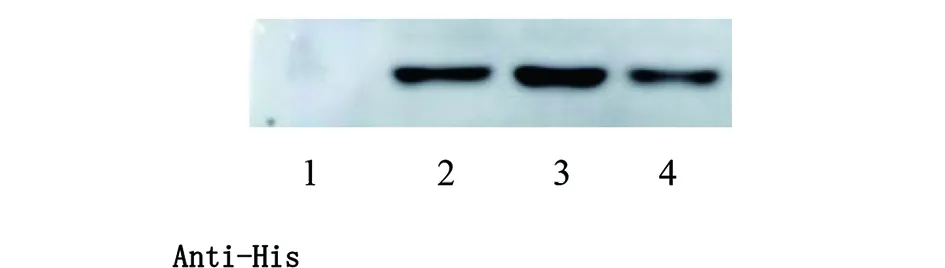
1:空載體;2:katG野生型蛋白;3:katG271突變型;4:katG315突變型。圖4 katG重組蛋白Western blot鑒定Fig.4 Identification of recombinant protein katG by Western blot
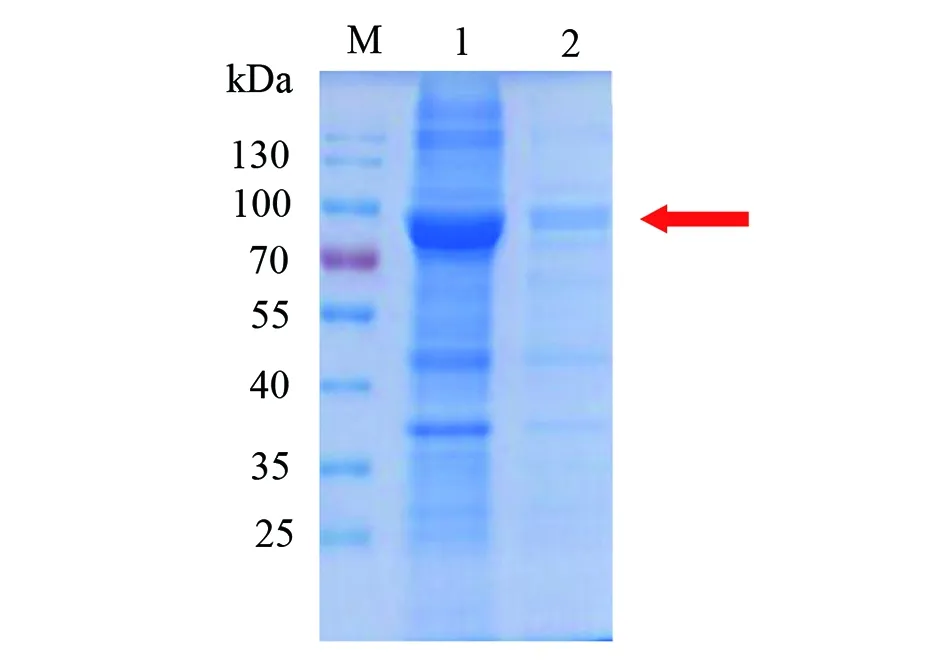
M: 蛋白分子質量標準;1:超聲破碎后上清;2:超聲破碎后沉淀。圖5 菌體超聲破碎后katG蛋白分布Fig.5 Distribution of katG protein after ultrasoniction
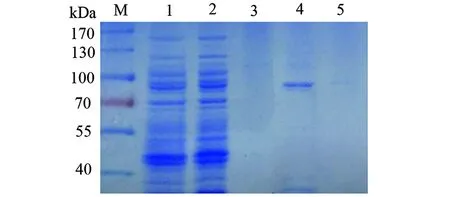
圖6 鎳親和層析純化的野生型katG蛋白SDS-PAGE電泳圖Fig.6 SDS-PAGE profile of wild-type katG purified by affinity chromatography
表1 katG重組蛋白的過氧化氫酶和過氧化物酶活性
Tab.1 Catalase and peroxidase activities of recombinant protein katGs
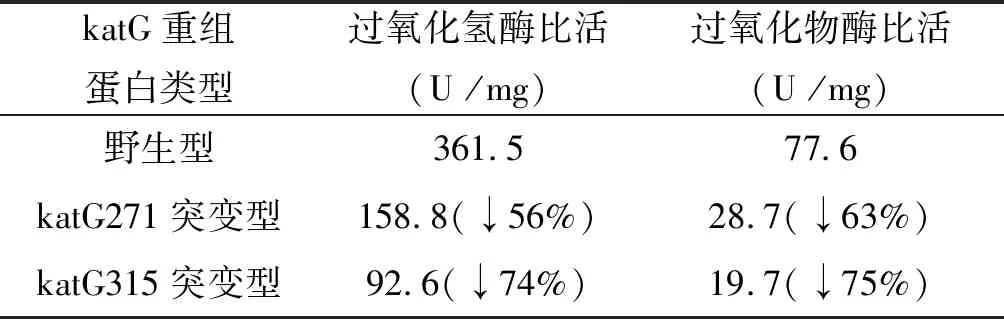
katG重組蛋白類型 過氧化氫酶比活(U /mg)過氧化物酶比活(U /mg)野生型361.577.6katG271突變型158.8(↓56%)28.7(↓63%)katG315突變型92.6(↓74%)19.7(↓75%)
注:1U過氧化氫酶為25 ℃、pH=7.0條件下,1 min內催化1 μmoL H2O2反應所需的酶量;1U過氧化物酶為25 ℃、pH7.0條件下,1 min內催化1 μmol鄰聯大茴香胺反應所需的酶量。
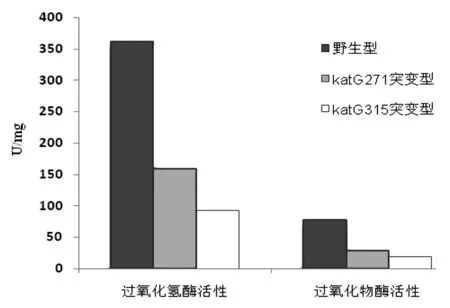
圖7 純化的katG蛋白過氧化氫酶和過氧化物酶活性比較Fig.7 Comparison of catalase and peroxidase activities of the purified protein katGs
2.2 分子對接
2.2.1異煙肼與katG蛋白活性空腔對接 圖8、9可以看出氧化反應的活性區域是蛋白質內部一個以卟啉為平面的狹長疏水區域,內部向外有一個狹小的通道口,約為10埃×10埃大小,僅允許體積較小的分子進入。圖中可以看出,異煙肼進入活性區域后,親水酰胺基團與ASP137殘基形成氫鍵,阻礙其它小分子進入。異煙肼的六元環為疏水基團,與卟啉環平行,兩者之間有疏水作用。分子對接發現,異煙肼均能進入野生型及其他突變體蛋白質的活性空腔,結合的位置相似。
2.2.2異煙肼與野生型katG蛋白對接 對接結果分析得,異煙肼與野生型katG蛋白結合的能量為-6.60 kcal/mol,可見反應是自發進行的。圖8可以看到,異煙肼與katG蛋白結合的位置位于氧化反應的活性中心,即卟啉環上方,酰胺氧分別與HIS108和TRP107氨基酸殘基分別形成2.3、2.2埃的氫鍵,結合在卟啉環上方。酰胺的氧位于Fe原子斜上方,距離為2.8埃。
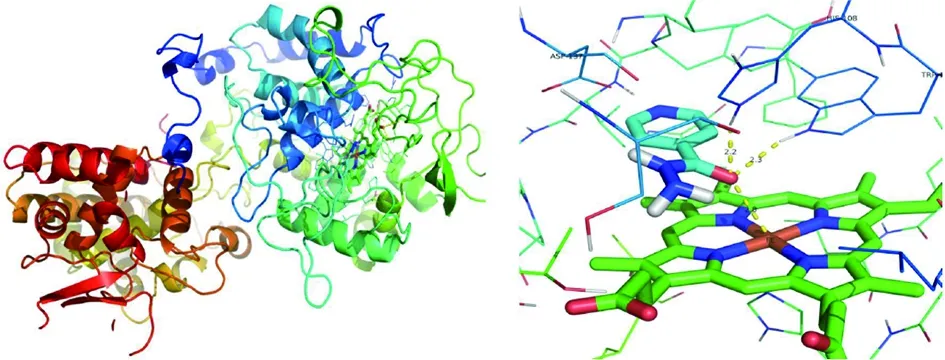
圖8 katG野生型與異煙肼結合示意圖Fig.8 Schematic diagram of the binding of katG wild type with isoniazid
簇分析發現,對接能量最低的10個構象都同屬唯一的簇,能量一致。可以看出,卟啉環上方的活性空腔既疏水又比較狹小,異煙肼含有親水的酰胺基團,無法進入口袋深部,只能結合在開口處,在其進入氧化活性空腔內后,占據了關鍵位置,阻礙其它分子的進入。
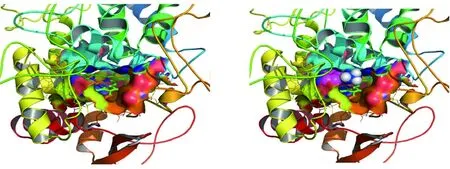
圖9 蛋白質氧化活性空腔示意圖Fig.9 Schematic diagram of the reactive cavity of protein oxidation
2.2.3異煙肼與Thr271Pro突變體katG蛋白對接 Thr271Pro突變體katG蛋白與異煙肼分子對接的結合自由能為-6.46 kcal/mol,較野生型升高,提示Thr271Pro突變體KatG蛋白與異煙肼結合的親和性降低。從圖10可以看出,酰胺的氧與Fe原子距離拉開到3.1埃,酰胺上的氫與ASP137上的氧形成了1.9埃氫鍵,而野生型蛋白與異煙肼沒有形成類似的氫鍵。此外,通過圖11發現ASP137殘基的羧基發生了一定角度的旋轉。
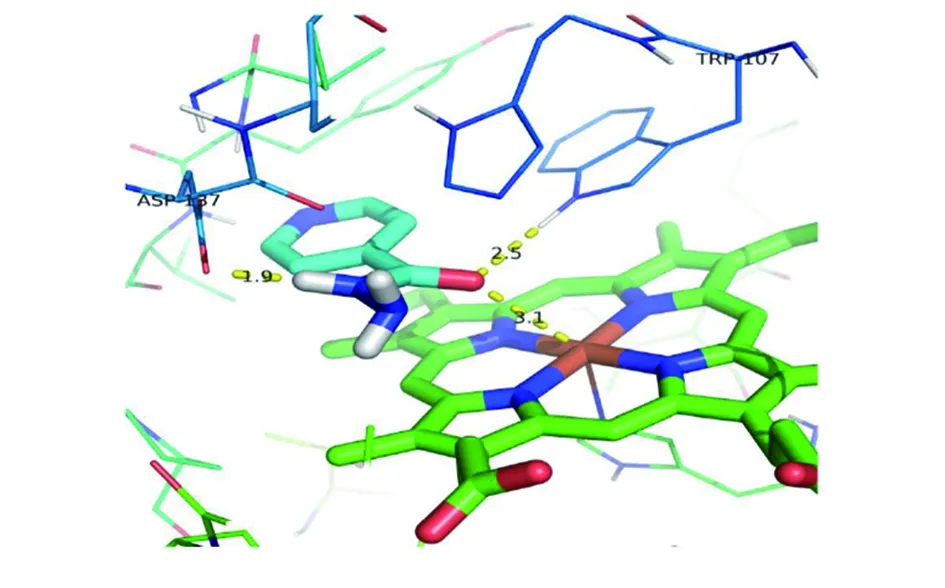
圖10 Thr271Pro突變體katG蛋白與異煙肼作用示意圖Fig.10 Schematic diagram of the interaction of katG(Thr271Pro) mutant with isoniazid
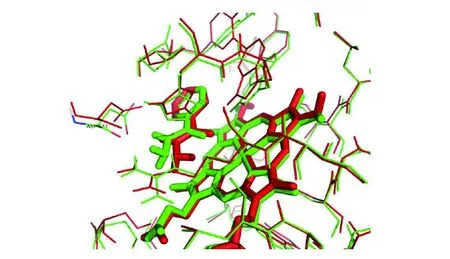
(紅色為katG野生型蛋白,綠色是katG Thr271Pro點突變蛋白)圖11 Thr271點突變蛋白與野生型蛋白結構疊加對比圖Fig.11 Comparison of structural superposition of Thr271Pro mutant protein and Wild-Type katG protein
3 討 論
1992年,Zhang等[6]研究證實katG基因缺陷的耐INH菌株獲得野生型katG基因后,恢復對INH的敏感性,因此認為katG基因在結核分枝桿菌INH耐藥機制中起著關鍵作用。進一步研究發現,不僅僅是katG基因的完全缺失,katG基因的點突變、堿基的插入與缺失都可能影響其編碼的過氧化氫-過氧化物酶活性,從而無法將INH轉化成殺菌活性形式的異煙酸,導致結核菌對INH耐藥[7]。但katG蛋白如何活化異煙肼及katG蛋白突變后導致異煙肼耐藥確切機制并不清楚。若能從基因型突變來判斷表型耐藥,將大大縮短藥敏培養所需時間,及時為臨床用藥提供參考。為此,研究人員在不同程度耐INH菌株中檢測出katG基因不同的突變位點,其中部分突變點被證實導致katG酶活性下降,認為可作為結核菌耐INH的重要標志[8-10]。然而,并非所有的基因位點突變都涉及結核分枝桿菌耐藥,有些為細菌進化自然多態性變化,有些卻為代償性突變。研究發現R463L位點的突變是常見的基因多態性,常常與katG基因其他突變位點一同發生,并且多見于異煙肼敏感菌株而不是異煙肼耐藥菌株[11]。
本實驗katG(Thr271Pro)突變結核菌株來自江西省異煙肼高水平耐藥株分離培養,MIC為1 mg/L,且katG(Thr271Pro)突變為katG基因唯一突變位點。而katG(AGC315ACC)突變為我國最為常見的基因突變形式[4]。本實驗構建katG野生型、katG(Thr271Pro)突變、katG(AGC315ACC)基因片段,重組至質粒pET-22b(+)。選擇該質粒原因是其具有氨芐青霉素抗性且能在katG蛋白C端加上6個His的融合標簽,轉化至大腸桿菌BL21(DE3)pLySs后,可在含氨芐青霉素培養基中挑選轉化陽性菌落進行下一步培養。對培養菌蛋白誘導并超聲破碎細菌后,最終可通過鎳親和層析方法純化出帶有6個His融合標簽的katG蛋白。酶活性測得:相比野生型(過氧化氫酶活性361.5 U/mg和過氧化物酶活性77.6 U/mg),katG271突變型過氧化氫酶和過氧化物酶活性分別下降56%和63%(158.8 U/mg和28.7 U/mg),katG315突變型過氧化氫酶和過氧化物酶活性分別下降74%和75%(96.2 U/mg和19.7 U/mg)。可以看出katG271突變型較野生型酶活性大幅度下降,但尚未達到katG315突變型下降程度。由此推斷該異煙肼耐藥菌株katG基因上唯一突變位點katG(Thr271Pro)突變引起了異煙肼耐藥。
通過分子模擬對接研究,能揭示耐藥靶點在結構層次上與藥物的相互作用,闡明藥物耐藥性的機理[12]。為探討katG(Thr271Pro)突變是如何導致異煙肼耐藥,進一步通過分子模擬對接技術發現katG突變酰胺的氧與Fe原子距離拉開到3.1埃,酰胺上的氫與ASP137上的氧形成了1.9埃氫鍵。野生型的蛋白與異煙肼結合沒有類似氫鍵,ASP137殘基的羧基發生了一定角度的旋轉。而且,Thr271Pro突變體KatG蛋白與異煙肼分子結合的親和性較野生型降低。從而可以推斷,271氨基酸殘基突變后,導致氧化反應活性空腔發生了一定的變形。
Brossier F等[13]在極高水平異煙肼耐藥菌株(>10 mg/L)中檢測到katG H270R突變,測得過氧化物酶-過氧化氫酶活性幾乎完全喪失,分子對接發現katG野生型270位置上組氨酸基團位于口袋近端,與亞鐵血紅素鐵離子形成配位鍵。270的突變形成空間阻礙,與亞鐵血紅素失去配位鍵連接,使亞鐵血紅素不能結合上katG蛋白。而本實驗研究對象271突變氧化反應活性空腔發生了一定的變形,并未完全與血紅素失去聯系,但酶活性表現非常低。吳海松等[14]在結核病異煙肼治療超過6個月患者體內檢測出270、271雙位點突變菌株,密碼子為(ACT-GTT),此體內研究表明該區域位點突變后,結核分枝桿菌仍在服用異煙肼患者中生長,進一步提示該位點與異煙肼耐藥極為相關。
研究結果提示:katG271位點及鄰近氨基酸殘基均為功能氨基酸,可能是維持katG蛋白活性的重要組分。其突變影響katG酶活性部分缺失及氧化反應活性空腔發生變形,與異煙肼耐藥關系相關。因此認為katG(Thr271Pro)可作為INH耐藥菌株檢測的參考標志,為臨床INH耐藥基因的篩查、開發基因芯片等提供重要理論依據。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
