NOP56在乳腺癌組織中的表達情況及對臨床預后的意義
2019-07-05 11:00:10林萍萍呂喜英吳練練李青山
生物信息學 2019年2期
曲 杰,林萍萍,呂喜英,吳練練,李青山*
(1.河北承德醫學院附屬醫院 腫瘤科,河北 承德 067000;2.武漢大學人民醫院 消化內科,武漢 430060)
乳腺癌是女性最常見的惡性腫瘤,發病率逐年升高,嚴重威脅女性生命健康[1]。NOP56 核糖核蛋白(NOP56 ribonucleoprotein, NOP56)是一種核仁蛋白,是C/D盒小核仁核糖核酸復合物(snoRNPs)的重要組成部分,在60S核糖體亞基組裝中發揮關鍵作用,涉及rRNA前體的早中期加工[2](MIM#614154)。研究報道,NOP56表達受原癌基因c-Myc調控[3],同時,NOP56也可調節c-Myc從而促進Burkitt淋巴瘤生長[4]。NOP56可以與hPygo2蛋白結合,促進人卵巢癌細胞生長[5]。在人乳腺癌細胞中,NOP56基因敲除后細胞集落形成能力和生長能力明顯降低[6]。然而,NOP56在人乳腺癌組織中的表達情況及臨床意義的大數據分析尚未見報道。
通過對美國生物技術信息中心(National Center for Biotechnology Information,NCBI)的基因表達綜合數據庫(Gene Expression Omnibus,GEO)[7]和美國國立健康中心(The National Institude of Health,NIH)的癌癥基因圖集數據庫(The Cancer Genome Atlas,TCGA)[8]公共數據集進行分析,探索NOP56在人乳腺癌組織中的表達及臨床病理關系,同時在K-M plotter 在線數據庫中研究NOP56的預后價值。進一步利用String、GSEA數據庫,挖掘乳腺癌中與NOP56存在互相作用的蛋白網絡,篩選與NOP56存在相互作用的基因并建立基因調控網絡,WebGestalt數據庫尋找靶向藥物[9],為乳腺癌的分子機制研究提供新的切入點和同時進一步探討臨床靶向藥物治療的分子機制。
1 材料與方法
1.1 數據資料收集
從NCBI的GEO數據庫下載原始數據,通過MAS5進行標準化,乳腺癌組織登錄號為GSE42568、GSE15852、GSE10780和GSE45255。從TCGA數據庫下載原始數據,通過RSEM標準化。納入的數據基本信息見表1。

表1 納入的4套乳腺癌基因芯片基本信息Table 1 Basic information of four GEO datasets of breast cancer
1.2 差異表達基因分析
在數據集GSE15852中,通過配對樣本T檢驗比較乳腺癌和配對癌旁組織NOP56表達差異。在數據集GSE42568和GSE10780中,通過非配對樣本T檢驗比較乳腺癌和正常組織NOP56表達差異;在TCGA數據庫中,通過非配對樣本T檢驗比較不同臨床病理分期的乳腺癌和配對癌旁組織NOP56表達差異;在TCGA數據庫中,通過非配對樣本T檢驗比較不同臨床病理分期乳腺癌組織和配對癌旁組織NOP56表達差異。
1.3 數據集篩選與臨床病理學參數相關性研究
TCGA數據集和GSE45255基因樣本量大且附帶臨床信息,利用此數據集進行進一步的相關性研究。其中臨床資料缺失的樣本被排除,根據表達譜數據,對樣本的NOP56表達進行由高到低排序,高于信號log2中位數的樣本作為高表達組,低于信號log2中位數的樣本作為低表達組。
1.4 Kaplan-Meier plotter在線數據庫分析
利用在線數據庫Kaplan-Meier plotter (http://www. kmplot.com/breast) 獲得的3 951例乳腺癌患者信息,以NOP56表達中位值(Median) 將樣本分為低表達和高表達組,利用缺省設置,在線分析NOP56對乳腺癌患者總生存期和無病生存期的預后價值。
1.5 NOP56互相作用基因篩選和功能富集分析
利用String(https://string-db.org)在線數據庫篩選人乳腺癌樣本中與NOP56存在互作關系的蛋白質并且獲得互作蛋白網絡圖。通過R3.5軟件計算互作基因在分子功能、生物過程、細胞組分以及三者平均的語義相似性并繪制熱圖。利用GSEA3.0軟件對上述互作基因進行富集分析。
1.6 靶向藥物富集分析
利用WebGestalt(http://www.webgestalt.org)在線數據庫富集分析互作基因的靶向藥物。
1.7 采用SPSS 20.0軟件進行統計學分析
生存分析采用Kaplan-Meier和log-rank檢驗法。臨床病理參數相關性分析,組間比較采用χ2檢驗及Fisher 確切概率法。以P< 0. 05 為差異有統計學意義。篩選共表達基因采用spearman和Pearson檢驗,spearman分數和 Pearson分數均大于0.3的基因被認為有共表達關系。功能分析采用Fisher精確檢驗,P<0.05的功能集合被認為差異有統計學意義。
2 結果分析
2.1 NOP56在人乳腺癌組織中高表達
在GEO數據庫的3個獨立數據集GSE42568、GSE15852和GSE10780中,與正常組織相比,NOP56在人乳腺癌組織中顯著高表達。在TCGA數據庫的數據集中,1 093例乳腺癌組織與112例正常組織相比,NOP56在人乳腺癌組織中顯著高表達(見圖1);不同臨床病理分期的乳腺癌組織與正常組織相比,NOP56均高表達(P< 0.001)(見表2)。
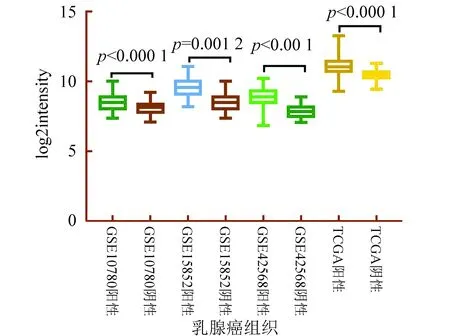
圖1 NOP56在人乳腺癌組織中高表達Fig.1 Overexpressed NOP56 in human breast cancer
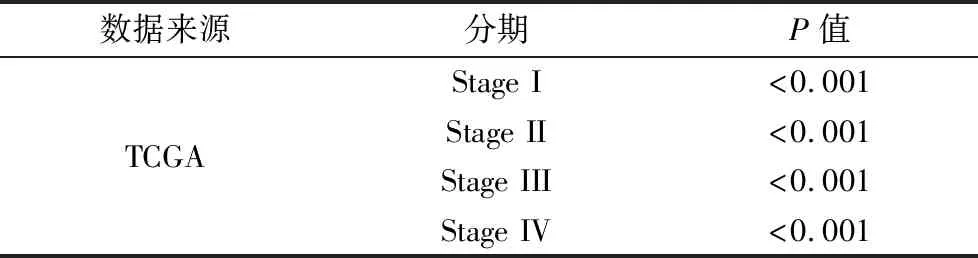
數據來源分期P值TCGAStage I<0.001Stage II<0.001Stage III<0.001Stage IV<0.001
2.2 NOP56高表達與患者不良臨床病理參數相關
2.2.1 分析GSE45255 數據集中年齡、淋巴結轉移、雌激素受體(ER)狀態、孕激素受體
(PRG)狀態、人表皮生長因子受體2(HER2)狀態、組織學分級和腫瘤大小等臨床病理參數與NOP56表達高低的相關性。結果顯示,NOP56 高表
達在組織學分級為G3的腫瘤樣本中更常見,而在分化程度較高的G1、G2腫瘤樣本中,NOP56低表達更多見(P< 0.001),提示NOP56高表達與乳腺癌的進展呈正相關。同時,NOP56與乳腺癌HER2狀態相關,在HER2狀態陽性的患者中高表達(P=0.019) 。NOP56的表達與年齡、淋巴結轉移、PRG狀態、ER狀態和腫瘤大小無顯著相關性(見表3)。
2.2.2 分析TCGA 數據集中年齡、淋巴結轉移、雌激素受體(ER)狀態、孕激素受體
(PRG)狀態、人表皮生長因子受體2(HER2)狀態和病理分期等臨床病理參數與NOP56表達高低的相關性。結果顯示, NOP56與乳腺癌HER2狀態、年齡、PRG狀態相關,在HER2狀態陽性的患者中高表達( P=0.019) ,在年齡< 55歲的患者中高表達(P<0.001),在PRG陰性的患者中高表達(P<0.000 1)。NOP56的表達與年齡、淋巴結轉移和ER狀態無顯著相關性(見表4)。GEO數據庫與TCGA數據庫結果一致。
2.3 NOP56高表達提示不良預后
K-M plotter生存分析顯示,NOP56高表達患者的無進展存活期(Disease free survival,DFS)和總存活期(Overall survival,OS)均明顯較低表達患者短(P< 0.001,見圖2)。這表明NOP56低表達與患者生存期縮短顯著相關,NOP56可能成為預測乳腺癌的預后指標(見圖2)。
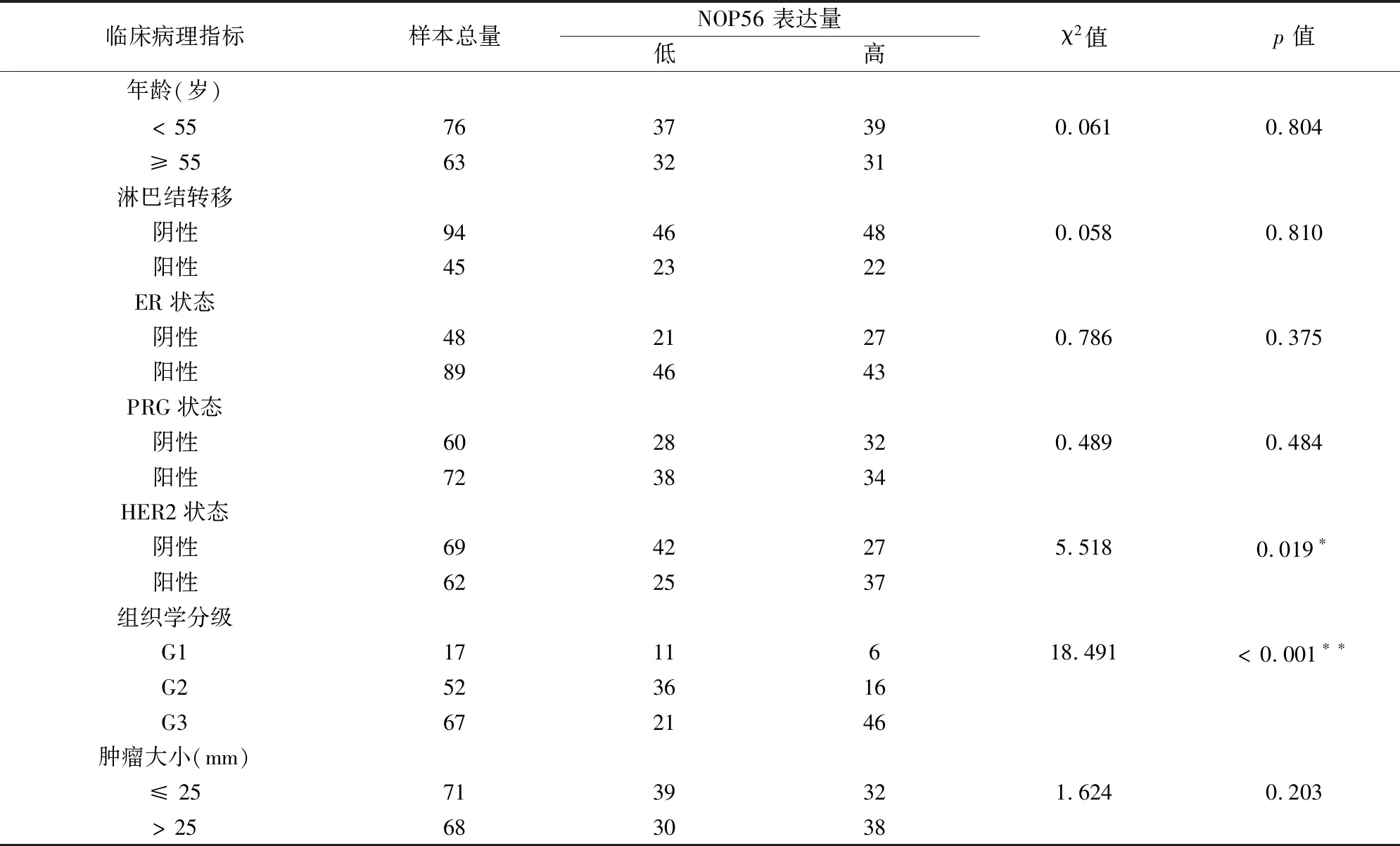
表3 不同NOP56表達與GEO數據庫139例乳腺癌患者臨床病理參數的關系Table 3 Correlation between NOP56 expression and clinicopathological parameters in 139 patients with breast cancer
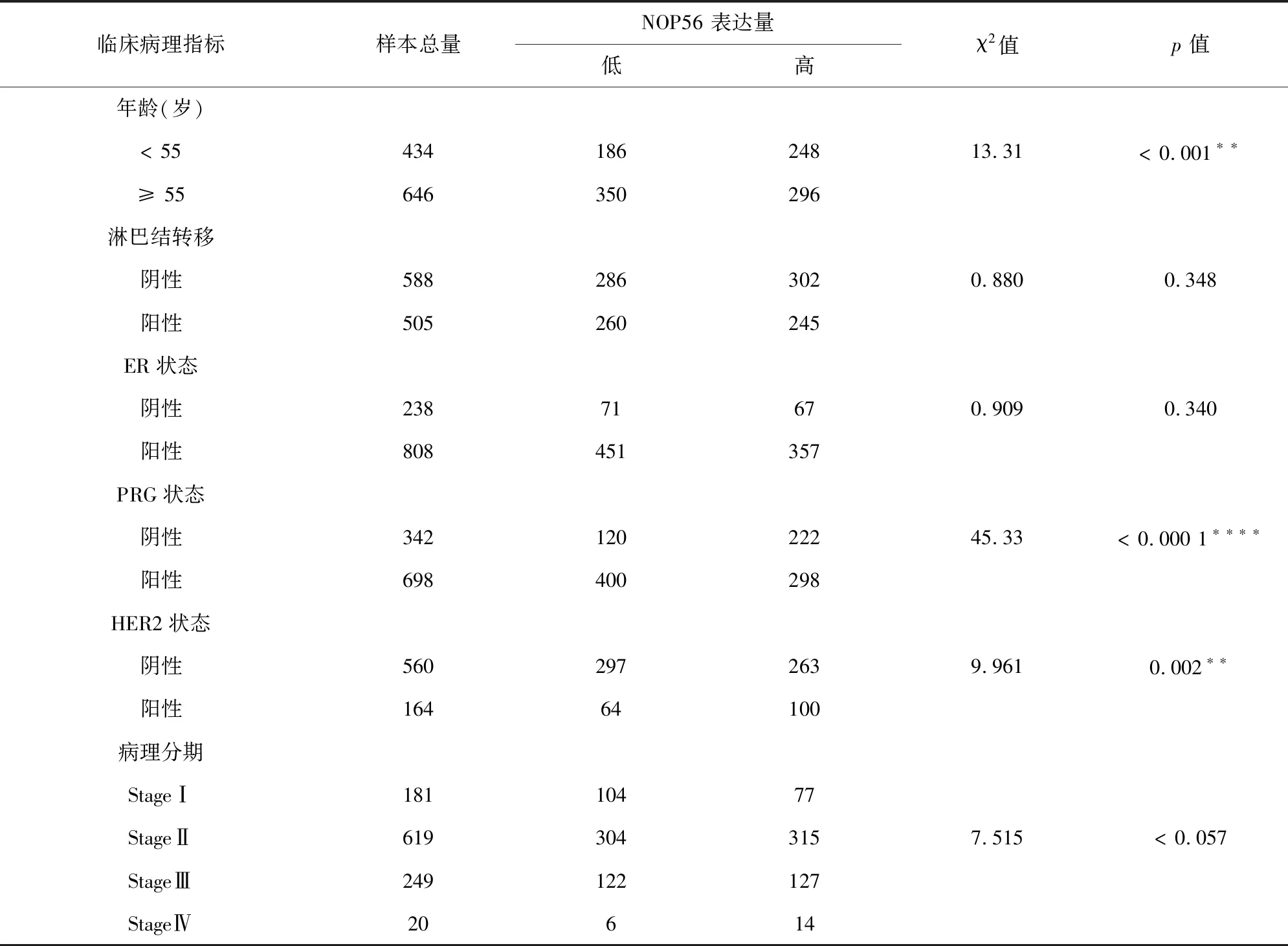
表4 不同NOP56表達與數據TCGA數據庫庫1 093例乳腺癌患者臨床病理參數的關系Table 4 Correlation between NOP56 expression and clinicopathological parameters in 1 093 patients with breast cancer in TCGA
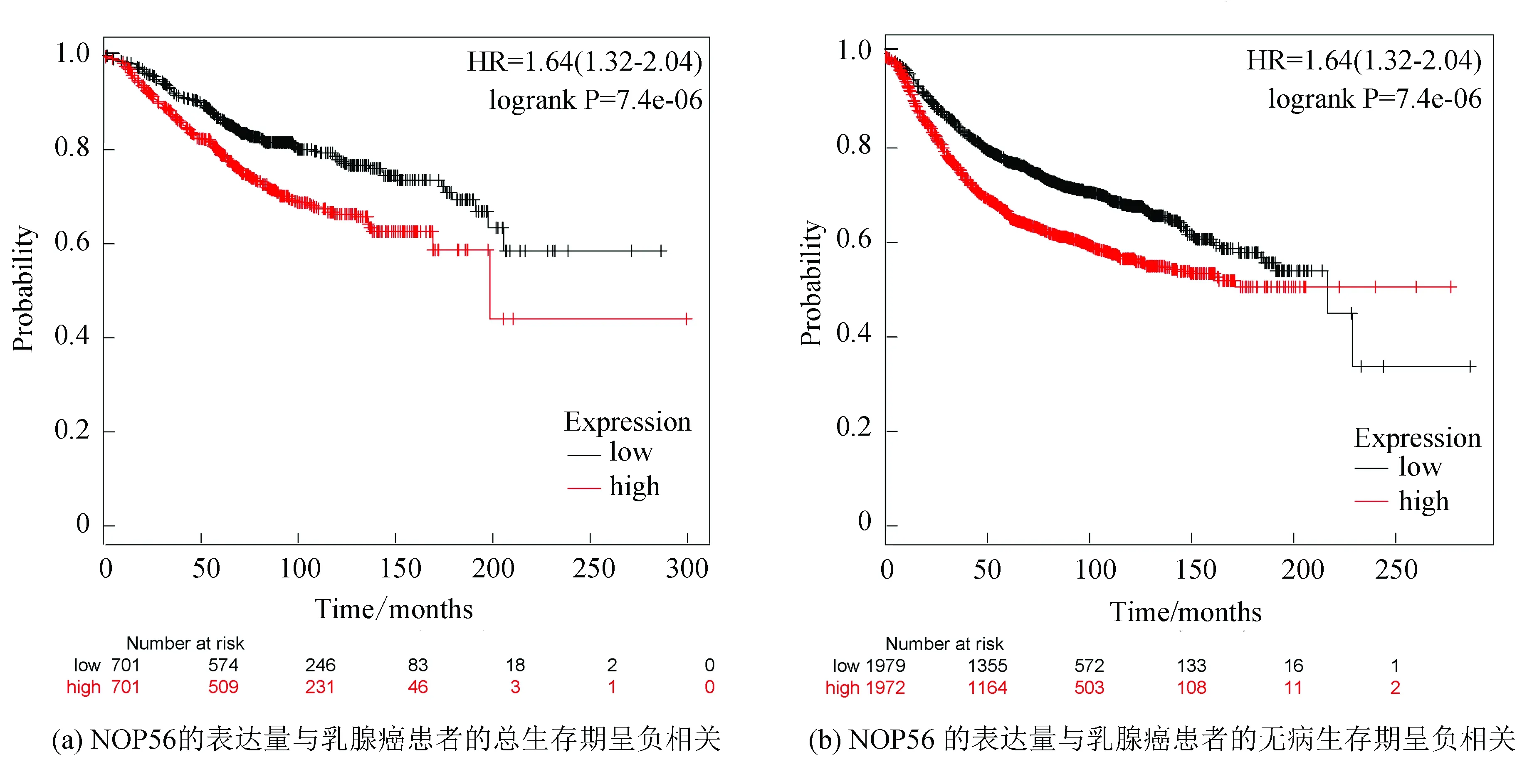
圖2 NOP56表達水平與乳腺癌患者生存時間關系Fig.2 Correlation between NOP56 expression and the prognosis of breast cancer patients
2.4 NOP56互作及其功能
String在線數據庫篩選人乳腺癌樣本中與 NOP56存在互作關系較強的基因10個。(見圖3(a)) R3.5進行語義相似性分析(見圖3(b)、(c)、(d)、(e))
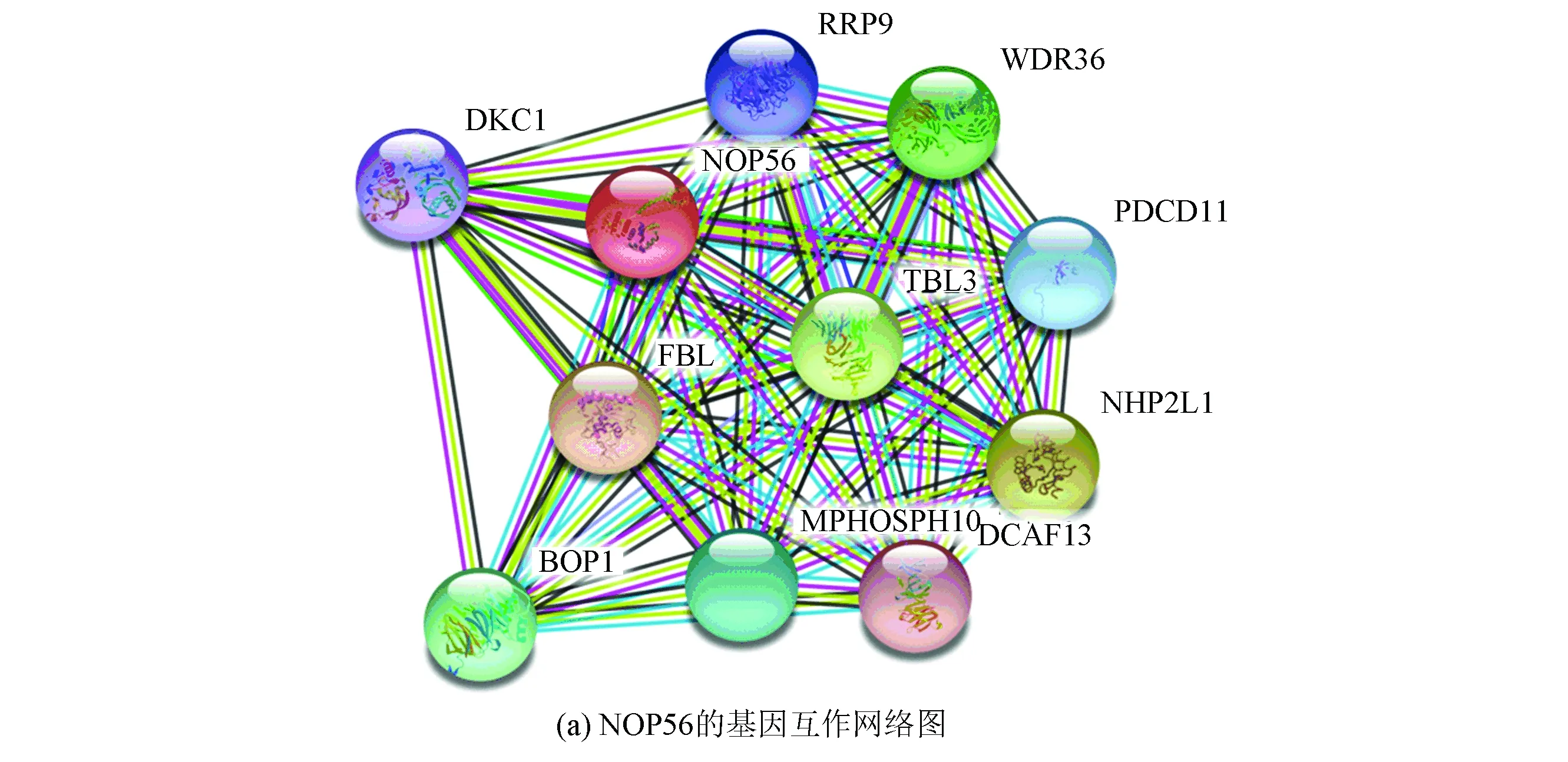
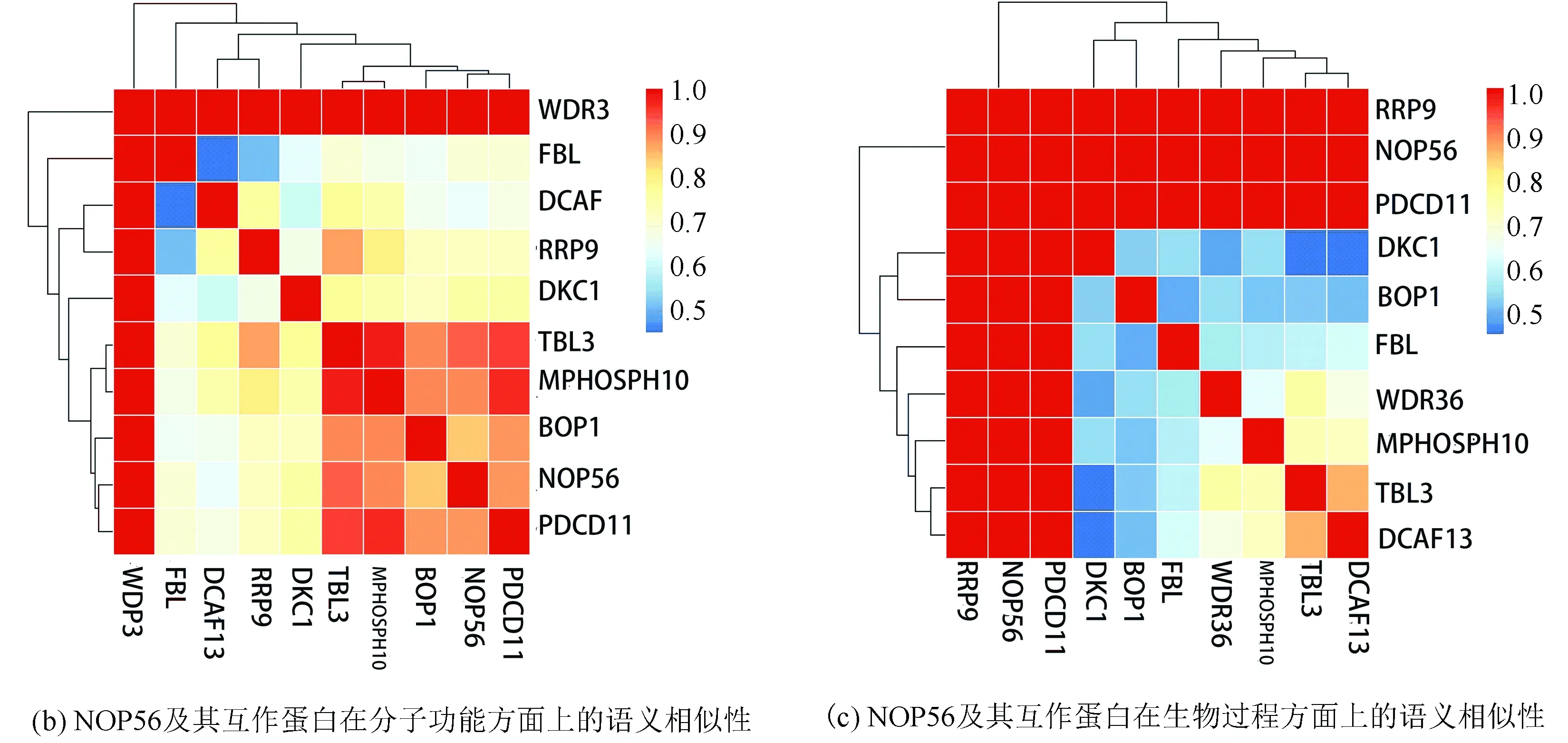
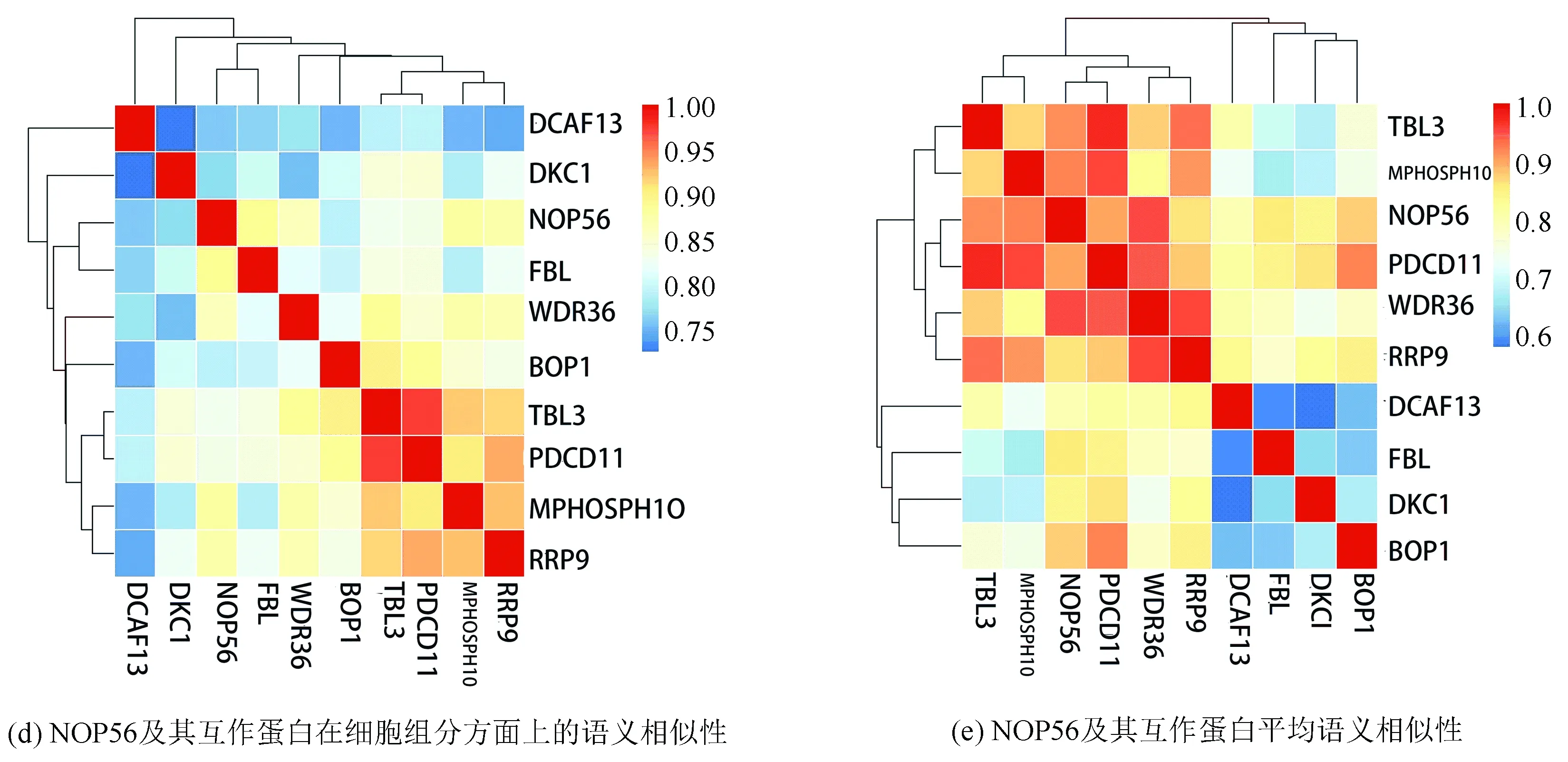
圖3 NOP56蛋白互作網絡以及共表達基因與NOP56分子功能、生物過程、細胞組分以及平均語義相似性Fig.3 Gene interaction network diagram of NOP56 and the molecular function,biological process,cellular component,and the mean semantic similarity between NOP56 and its co-expressed genes
利用GSEA數據庫對互作基因進行富集分析,結果顯示這些基因主要富集在RNA加工,細胞分裂和DNA復制等與腫瘤發生發展關系密切的功能上(見圖4)。
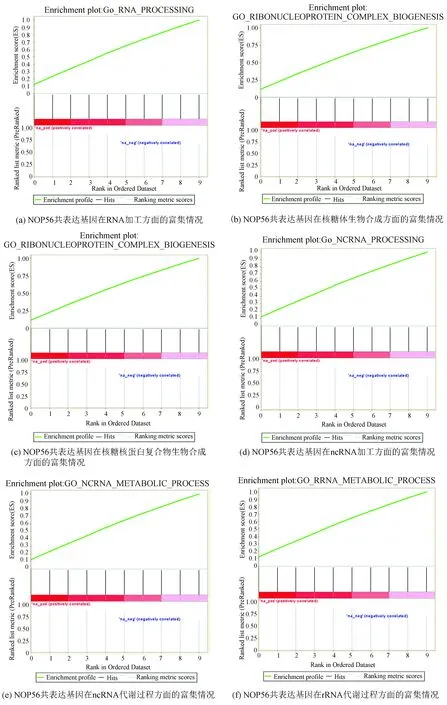
圖4 NOP56共表達基因主要富集在RNA加工和細胞分裂等與腫瘤發生發展關系密切的功能上Fig.4 NOP56 co-expression genes are mainly enriched in the functions closely related to the tumor development such as RNA processing and cell division
2.5 NOP56互作基因靶向藥物
WebGestalt網站分析顯示,以上互作基因靶向藥物有5種,其中互作基因作為靶基因最多的為放線菌素D(見表5)

表5 靶向基因藥物Table 5 Targeted gene drugs
3 討 論
乳腺癌的發生發展離不開異常的蛋白質生物合成[6],而細胞中蛋白質的合成需求與核糖體生物合成活性相協調,當某些關鍵的核糖體蛋白水平上調后,可通過調節其他蛋白的表達影響細胞生長代謝[10]。研究表明,核糖體生物合成速率增加促進原癌基因Myc表達,增加癌細胞增殖能力[8],同時可通過釋放MDM2蛋白降解抑癌基因p53[12]和pRb[13],增加癌癥患病風險。核糖體生物合成活躍可促進乳腺癌細胞侵襲能力,增加乳腺癌惡性程度[14]。NOP56是小分子核仁核糖核蛋白(snoRNP)復合體的核心蛋白質成員,直接對rRNA前體核糖部分進行甲基化修飾,在核糖體生物合成中發揮重要作用[15]。NOP56突變可導致60S核糖體亞基裝配缺陷,rRNA加工受抑[15]。研究表明,NOP56可調節原癌基因c-Myc促進Burkitt淋巴瘤細胞生長[4],同時可以與hPygo2蛋白結合,促進卵巢癌細胞生長[5]。本研究通過生物信息學分析,首次在人乳腺癌組織中發現NOP56顯著高表達,且NOP56高表達與乳腺癌不良臨床病理參數和預后相關。為進一步揭示其作用機制,我們探索了乳腺癌中NOP56共表達基因、互作蛋白及其功能,并構建蛋白互作網絡圖。
通過GEO和TCGA數據庫,我們首次發現NOP56在人乳腺癌中顯著高表達。在GSE45255中,數據顯示NOP56高表達患者乳腺癌分化程度低。這與已有的研究一致,Su H等人發現NOP56促進人乳腺癌細胞集落形成能力和生長能力[6]。此外,我們發現NOP56在HER2陽性患者中高表達。HER2過表達與乳腺癌的發生、發展、侵襲和轉移密切相關[16]。HER2基因編碼酪氨酸激酶活性跨膜糖蛋白,與相應配體結合導致胞內酪氨酸激酶磷酸化,引起細胞生長、增殖相關信號通路如NF-κB等的連鎖反應,促進細胞增殖分化[17]。HER2陽性乳腺癌細胞c-Myc表達水平高,敲低HER2基因后c-Myc表達降低[18],而c-Myc可直接促進NOP56表達[3]。這說明乳腺癌中HER2可能通過激活細胞生長增殖相關信號通路上調c-Myc等原癌基因從而促進NOP56表達。我們進一步利用在線數據庫K-M plotter,對3 951例乳腺癌患者進行預后分析,發現NOP56高表達乳腺癌患者生存期顯著降低。乳腺癌組織中NOP56表達水平,或許可以作為乳腺癌患者新的檢測指標和遠期預后評價指標,但還需大樣本的臨床數據進一步驗證。
為了進一步理解乳腺癌中NOP56異常表達的機制,篩選出NOP56最顯著的互作基因10個。單一的基因異常往往伴隨下游基因的連鎖異常反應,這是多種癌癥相關基因異常促進癌癥發展的重要原因之一。功能分析顯示,NOP56共表達基因主要參與RNA加工,細胞分裂和DNA復制等與腫瘤發生發展關系密切的功能。這些結果與現有的實驗結果一致,Victoria H.等[5]發現NOP56 基因過表達后,Burkitt淋巴瘤細胞的增殖明顯加快;Yuichi J.等[19]發現在乳腺癌上皮細胞中Nop56促進細胞周期G1期縮短,是細胞周期進程的重要介質。研究證明NOP56共表達網絡的準確性,提示NOP56可能通過影響這些共表達基因來發揮其促癌作用。
最后,我們篩選出了NOP56互作基因富集的靶向藥物,選取了富集基因最多的放線菌素類中的放線菌素D。放線菌素D(更生霉素)是第一種顯示出具有抗腫瘤活性的抗生素,是臨床上廣泛使用的抗腫瘤藥物,現在常用于各種小兒腫瘤,如腎母細胞瘤,橫紋肌肉瘤和尤因氏肉瘤的治療[20-22]。放線菌素D是一種DNA嵌入劑,它更偏向于富含GC的DNA序列[ 23 ]。Perry RP等人發現低濃度的放線菌素D可以破壞核糖體正常的生物過程,最終導致細胞周期停滯和細胞凋亡[ 23-25]。Das T等人研究發現放線菌素D可以下調SOX2表達,從而誘導乳腺癌干細胞死亡[26]。通過這些研究我們可以假設,放線菌素D通過靶向作用于NOP56以及其互作基因達到對乳腺癌的治療作用,放線菌素D的治療作用可能與NOP56的表達量相關,而這些假設還需要進一步的實驗以及臨床驗證。
綜上所述,利用公共數據庫證明乳腺癌組織中NOP56高表達,NOP56促進腫瘤分化,在HER陽性患者中表達量高,且與乳腺癌患者的不良預后密切相關。機制上,我們研究了乳腺癌中NOP56共表達基因,其功能主要為RNA加工,細胞分裂和DNA復制等與腫瘤發生發展關系密切的功能,并構建基因互作網絡圖,發現互作基因靶向藥物明顯富集在放線菌素D上。系統地研究了NOP56在乳腺癌中的表達與臨床預后、臨床病理狀態以及組織學分級的關系,探討了分子機制以及與臨床靶向藥物的關系,為乳腺癌防治提供潛在的新的預測指標,完善了臨床靶向藥物使用的分子機制。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
