血栓病治療藥物-纖溶酶的生物來源
2019-07-05 09:43:16史延茂
生物信息學 2019年2期
董 超,馬 萱,史延茂
(河北省科學院 生物研究所, 石家莊 050081)
隨著人們生活水平提高、飲食結構變化和環境惡化等因素的影響,目前血栓性心腦血管性疾病已經嚴重危害人類的健康,不但發病率逐年增加 ,而且是致殘率和死亡率最高的疾病之一,所以醫學行業對相關血栓病的治療和預防藥物一直非常重視。人體一般正常情況下,凝血系統、抗凝血系統及纖溶系統在體內共同作用,保持動態平衡。當血液中凝血系統被不適當激活、或凝血-抗凝血體系中任一環節出現調節障礙,就容易引發血栓形成[1]。血栓藥物對于血栓栓塞性疾病的預防與治療,其臨床用藥作用機制主要分為抗血小板凝集、抗凝和溶栓三大類,其中前兩種是預防性的,后一種是治療已經發作的血栓。
血小板凝集是正常凝血中的一個關鍵環節,血小板的粘附、聚集和釋放反應是傷口止血的正常過程,由于是級聯激活,所以簡稱“凝血瀑布”。動脈粥樣硬化斑塊破裂、嚴重感染和分娩有時也會開啟這個過程,引起心腦梗、彌漫性凝血和羊水栓塞。從血小板發生聚集作用的機制來看,抗血小板凝集藥主要分為四類:(1)抑制環氧合酶和花生四烯酸代謝產物的藥物,如口服的阿司匹林;(2)增加血小板內CAMP含量的藥物,如潘生丁等;(3)作用于血小板膜特異激動劑和受體的藥物,如氯吡格雷;(4) 抑制血小板活化因子(PAF)藥物,如銀杏葉等。
抗凝藥是指能降低機體的凝血功能、防止血栓形成或進一步發展的藥物。在多種生理及病理狀態下,人體凝血活性可顯著增強, 如房顫患者的血液瘀滯,表現為某些凝血因子水平升高或活性增加,而高凝狀態是血栓性疾病的發病基礎。常用的抗凝藥物注射類有肝素,口服的有華法林、利伐沙班等,還包括重組水蛭素衍生物和L-精氨酸衍生物。一般都是作用于凝血過程中的凝血因子,控制凝血過程,而不是針對已經形成的血栓。
纖溶系統是降解已經形成的血栓的體系(見圖1),包括纖溶酶原、組織型纖溶酶原激活物、尿激酶型纖溶酶原激活物、纖溶酶原激活物抑制劑幾種物質。纖溶酶原在纖溶酶原激活劑的作用下,變成有活性的纖溶酶,纖溶酶作用于纖維蛋白凝塊,從精氨酸-賴氨酸鍵位置分解,得到可溶性的纖維蛋白降解產物,以保持血管通暢,防止血栓進一步擴大。

圖1 纖溶酶降解纖維蛋白示意圖Fig.1 Schematic diagram of the degradation of fibrous protein by fibrinolytic enzyme
抗血小板和抗凝血兩類藥物只能抑制或者預防血栓的形成,而不能從根本上溶解血栓[2-3]。溶栓藥物主要用于已形成的血栓,因此溶栓治療是目前臨床界治療血栓性疾病最為重要、有效且可靠的手段。纖溶酶作為治療血栓性疾病的主要溶栓藥物,一直倍受藥物開發者的高度關注。科研人員在人體、動物、植物和微生物中陸續發現了多種具有溶栓活性的成分,有些已經在臨床上使用,有些纖溶酶的基因序列進行了生物學的改造,這為尋找新型、高效的溶栓藥物提供了更多的思路,本文主要針對國內外發表的纖溶酶生物來源和特性進行了總結,并對產纖溶酶的微生物種群進行了分類比較。
1 來源于人體組織的溶栓藥物
溶栓酶可來源于人體,比如人體內的子宮、腎臟、肺組織,以及癌細胞培養組織,都可以產生纖溶酶,其中開發最早、處理工藝最成熟的是來源于健康人尿液的尿激酶 (UK),后來采用細胞培養開發了尿激酶纖溶酶原激活劑(uPA)和組織纖溶酶原激活劑 (t-PA)等。
1.1 尿激酶(Urokinase ,UK)
尿激酶是從健康人尿或腎組織培養液中獲得的絲氨酸蛋白酶,有兩種,分子量分別是33 KD和54 KD。尿激酶可以直接作用于纖維蛋白凝塊,也可以影響內源性纖維蛋白溶解系統,催化裂解纖溶酶原成纖溶酶,從而發揮溶栓作用。尿激酶溶栓能力強且毒性很低,臨床使用過程中也罕見過敏反應,是現在臨床治療急性血栓的主要藥物之一。但是尿激酶缺乏溶栓特異性,有導致出血的危險,臨床應用要嚴密的監護。目前醫院臨床還在為急性血栓患者使用,國內廠家有北京賽生藥業有限公司。
1.2 尿激酶纖溶酶原激活劑(uPA)和組織纖溶酶原激活劑 (t-PA)
尿激酶型纖溶酶原激活劑是尿激酶的前體,是一種絲氨酸蛋白水解酶,主要由健康人尿、血液或條件培養液中提取或者基因重組得到,可激活纖溶酶原轉變為纖溶酶,同時能夠降解血栓中的水不溶纖維蛋白形成水溶性降解片斷,從而使血栓溶解。組織型纖溶酶原激活劑,又稱纖溶酶原激活因子(Tissue plasminogen activator),屬于絲氨酸蛋白酶類,可特異性水解纖溶酶原中精氨酸-賴氨酸之間的肽鍵,從而能使纖溶酶原轉變為纖溶酶。這兩種藥物溶栓特異性較強,極大的減小了因系統纖溶活性導致的出血,同時還具有抗血小板凝集的作用,但是體內半衰期都較短[4]。臨床藥物有阿替普酶和瑞替普酶,生產技術一般采取基因克隆細胞培養,工藝和設備要求比較高,所以市場價格一直不低。
2 來源于動物組織的溶栓藥物
2.1 蚓激酶(Vermis kinase)
蚯蚓在藥學上稱地龍,蚓激酶是日本的Mihara首先從特種蚯蚓中提取的一組蛋白水解酶,分子量23~30 KD,具有纖溶和抗凝的作用[5]。蚓激酶可以直接降解纖維蛋白,也可以激活纖溶酶原,具有直接和間接溶栓功能;也可吸附凝血因子,并將其水解,具有間接抗凝的作用。對于蚓激酶的安全性,Tjandrawinata等[6]對于蚓激酶DLBS1033口服劑量進行了臨床安全性評價,研究表明DLBS1033以每日490 mg的劑量分3次給予,在健康成人中是安全的。這說明蚓激酶具有良好的生物安全性,值得深入研究開發[7]。
蚓激酶是一組群蛋白的總稱,有些科研人員進行了分離純化的研究,到目前報道的蚓激酶成分超過20種,并且口服蚓激酶具有溶栓和抗凝的效果,所以推測一般的蚓激酶提取物除了蛋白成分復雜,還可能具有糖類或其他小肽類成分,可以抵抗胃酸環境對蚓激酶的影響。國內口服蚓激酶產品于1992年上市,如國內的百奧、博洛克、普恩復和降寧蚓激酶都采用腸溶膠囊。蚓激酶口服的效果雖然出血性小,費用低,但是作用相對較慢,靜脈或注射的針劑開發是其發展方向之一。
2.2 蛇毒抗栓酶類
李時珍的《本草綱目》就有蝮蛇能治療半身枯死的敘述。上世紀60年代國外從蝮蛇毒液中提取到了有纖溶活性的酶,80年代國內開始提取研究,命名為降纖酶或降纖酶(Defibrinogenase),并且于1995年頒布了國家標準[8]。蛇毒抗栓酶能直接溶解血栓纖維蛋白和纖維蛋白原,從而降低血液黏度,同時可抑制血小板聚集。因而具有溶栓、去纖和抗凝的作用,不良反應有出血傾向和其它臟器的受損可能性。日本的東菱精純抗栓是從巴西矛頭蛇毒液中提取,可以用于急性腦梗和閉塞性缺血;2001年曹郁等將TSV-PV基因插入供體質粒pFastBacHTa中并在Tn-5B1-4細胞中表達,經過Western及酶活檢測表明,TSV-PV蛋白在昆蟲細胞內的表達具有較高的活性,為天然溶栓藥物的制備提供了新來源[9]。蛇毒抗栓酶由于來源特殊,多采用基因重組的方法制備。由于制備工藝不一,質量控制有難度,并且對其它臟器具有不良作用較多,所以臨床使用一般用于急癥或重癥病例,少于其它同類溶栓藥物的應用。
2.3 吸血蝙蝠唾液纖溶酶原激活劑
吸血蝙蝠唾液纖溶酶原激活劑最早是美國西點軍校研究室從南美吸血蝙蝠唾液中分離的,一種能與纖維蛋白特異性結合的高分子蛋白(Bat-PA),其溶栓能力與組織纖溶酶原激活劑相同,但纖維蛋白的特異性較高且免疫原性極低[10]。吸血蝙蝠唾液纖溶酶原激活劑半衰期較長,用量少,但由于吸血蝙蝠是稀有物種,其消耗性研究受到了嚴格的控制,因此重組蛋白是其應用開發的主要研究方向。Ma等[11]在吸血蝙蝠唾液中鑒定出一種新型溶栓酶Desmolaris,在抗血栓形成中起重要作用,同時能夠對多巴胺引起的血管通透性增加有緩解作用,是一種新型的靶向抗凝血劑。西南大學的劉堰和蘇暢等進行了DSPAa1在畢赤酵母中的表達研究[12],但是目前還是處于實驗室階段。
2.4 水蛭素(Hirudin)
早在17世紀歐洲醫學工作者就發現了水蛭的抗凝血作用,成分分離后命名為水蛭素,含有65-66個氨基酸的多肽鏈,分子量約7 KD。水蛭素與凝血酶可以形成非共價可逆性復合物,是目前最安全的天然凝血酶抑制劑[13]。但是天然水蛭素來源有限,所以現在大多采用基因重組的手段獲得。1986年RP Harvey首次在大腸桿菌中表達,獲得重組水蛭素(r-HVZ);國內的譚樹華等在水蛭素基因克隆方面研究較早,并且在畢赤酵母和乳酸菌表達水蛭素成功[14]。現在關于水蛭素的研究主要在鼻腔給藥方面,因為水蛭素口服效果吸收很差。
2.5 來源于動物的纖溶酶分類比較
纖溶酶可來源于動物,作用方式不盡相同,且各有優勢和不足,其對比見表1。
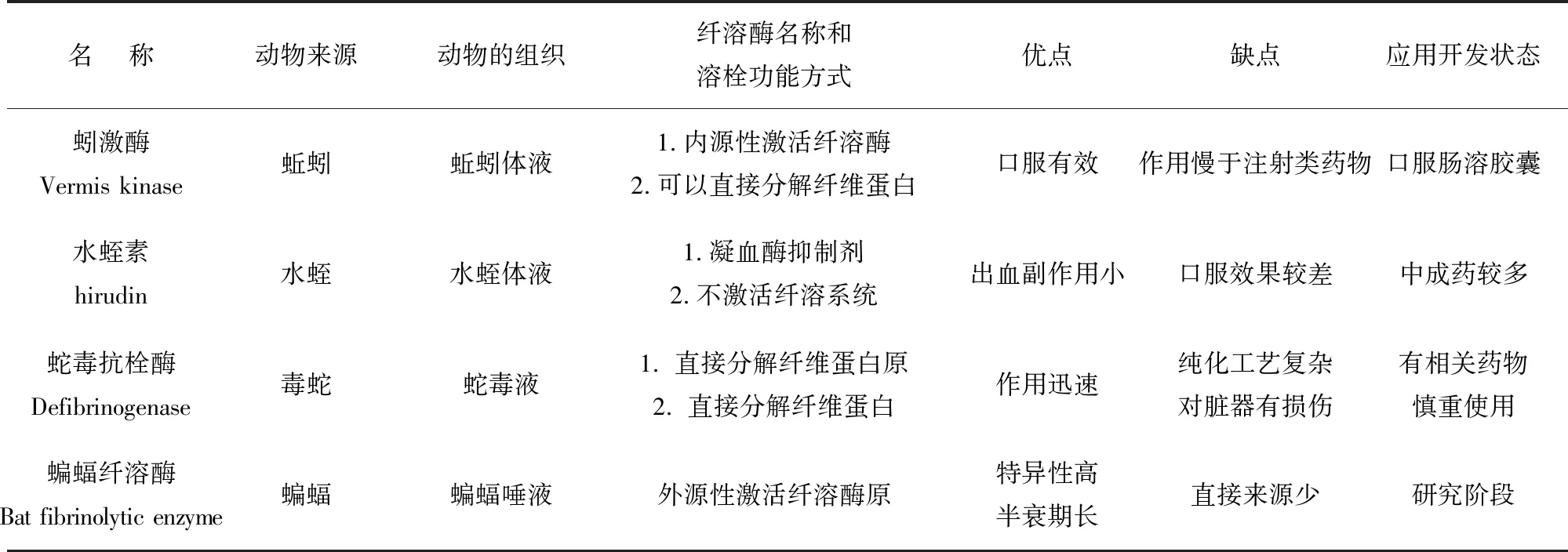
表1 產纖溶酶的動物種類Table 1 Animal types of fibrinolytic enzymes
3 來源于微生物組織的溶栓酶
微生物種類繁多,生長繁殖快,而且易于培養,生長條件較易控制,是溶栓藥物的重要來源。作用機理不同微生物來源的溶栓藥物可分為兩類:(1)纖溶酶原激活劑,如鏈激酶 (SK)、葡激酶 (SaK),其作用原理是將纖溶酶原激活成為纖溶酶,纖溶酶再降解構成血栓骨架的纖維蛋白,從而溶解血栓;(2)纖溶酶類物質,可以直接降解血栓中的纖維蛋白,如納豆激酶 (NK),以及來源于芽孢桿菌、糞鏈球菌、鏈酶菌等的纖溶活性物質。當前微生物溶栓藥物不僅可以從自然界中篩選,還可以通過改造的基因工程菌,提高生產效率和溶栓作用的專一性。
3.1 鏈激酶 (Streptokinase, SK)
鏈激酶是于1949年發現的由β-溶血鏈球菌(Streptococcushomolyticus,也稱乙型溶血鏈球菌)分泌的一種胞外酶,是最早應用于臨床的纖溶酶原激活劑,分子量約47~50 KD。鏈激酶進入血液后,須與纖溶酶原結合,使纖溶酶原活化位點暴露,從而使纖溶酶原成為有活性的纖溶酶。纖溶酶可以迅速降解纖維蛋白原成小分子產物,使這些降解產物不能參與血纖維網的形成過程,從而阻礙血栓形成;同時纖溶酶可以通過降解纖維蛋白,使血栓溶解[15]。經臨床應用表明,鏈激酶高效且價廉,但是半衰期短,容易引起過敏和抗原性抗體反應。為了使鏈激酶的應用更具安全性和有效性,現在使用基因重組的方法制備SK,取得一定的進展。
3.2 葡激酶(Staphylokinase, SaK)
葡激酶是由金黃色葡萄球菌分泌的一種胞外蛋白質水解酶,分子量為16.5 KD。葡激酶的作用機制與鏈激酶相似,也是一種間接纖溶酶原激活物。SaK具有極強的纖維蛋白特異性,且臨床使用過程中未見過敏反應。但由于葡激酶來源于細菌,仍具有一定的潛在免疫原性,最初的動物實驗發現出血傾向和毒副作用較大,造成對其研究一度沉寂。隨著基因工程的發展,克隆Sak基因在大腸桿菌中獲得高效表達,重組SaK特異性高易于大量制備,且無變態反應,價格相比tPA低廉許多。
3.3 納豆激酶(Nattokinase,NK)
納豆激酶是1987年日本的Sumi首先在納豆(日本的一種傳統食品)中發現的一種具有溶栓活性的絲氨酸蛋白酶,是一種起步晚但發展迅速的溶栓劑。納豆激酶不僅可以直接溶解血栓,而且可激活血漿中的纖溶酶原。納豆激酶的纖維蛋白原水解活性遠低于纖溶酶和尿激酶,這表明納豆激酶在發揮纖溶作用的同時,不水解血漿蛋白原,因此不易引起出血。納豆激酶生產成本低,溶栓能力強,半衰期長,安全性高,具而有很大的開發價值。國內研究納豆激酶較早的是東北農業大學的徐仲等,測定了濕納豆的酶活性是1 600 IU/g;南京工業大學的熊曉輝等[16]和河北省科學院生物研究所的董超等[17]通過優化發酵技術,酶活水平都達到了2 200 IU/ml以上;北京大學基礎醫學院韓慧明,李詠梅已通過構建工程菌使納豆激酶基因在大腸桿菌中表達[18]。但是到目前為止,尚未有關于納豆激酶的注冊藥物上市,大多以保健食品形式銷售。
3.4 鏈霉菌產生的新型纖溶酶
以放線菌產纖溶酶的報道很少,鏈霉菌屬的一些菌株可以產生絲氨酸蛋白酶,因此利用鏈霉菌產生的新型纖溶酶來制備溶栓藥物可能是一種新的嘗試。
鏈霉菌是以產抗生素引起人們關注,但是關于分泌纖溶酶的研究不是太多。1996年法國血液病研究所的F.bono等從一株鏈霉菌中得到纖溶酶,分子量約33 KD,等電點pH 6.7;印度的Chitte和Ratnakar等從高溫溫泉中分離得到的鏈霉菌SD5可以分泌一種耐高溫的具有溶栓作用的蛋白酶。該酶可以特異性地裂解纖溶酶原的肽鍵使其轉變為纖溶酶。該酶在37~60 ℃和pH值6.0~9.0時穩定性較高,由于范圍比較寬,有可能規模生產的成本較低[19-20]。 朝鮮大學藥學院P.Mander等從鏈霉菌CS624和684中分別得到了分子量18 KD和35 KD的纖溶酶,并且后一種還可以耐受高溫,在45 ℃酶活最高,確定是絲氨酸蛋白酶[21-22]。
1999年中國醫學科學院的王駿等從云南土壤篩選得到的鏈霉菌 Y405可產多種纖溶活性物質,其中有一種新型具有纖溶活性的蛋白酶-SW1,經研究表明,它不是一種纖溶酶原激活劑,而可以直接降解血栓中的纖維蛋白,效果與同劑量的尿激酶相當[23]。2001年中國醫學科學院和中國協和醫科大學龔勇等從鏈霉菌C3662中分離出一種分子量約為30 KD的具有纖溶活性的蛋白酶,并使用鳥槍法對該纖溶酶基因進行克隆,實驗結果表明該基因為一種新的基因[24]。河北工業大學的侯正欣等從海洋環境中篩選了一株產纖溶酶的鏈霉菌,發酵水平可以達到2 190 IU/ml,但是不具有激活纖溶酶原的作用[25]。
3.5 假單胞菌產生的纖溶酶
假單胞菌屬是指一群無芽孢、無莢膜的不能發酵糖類的革蘭氏陰性桿菌,專性需氧,廣泛分布于土壤、水和空氣中,是人類或動物的條件致病菌。關于假單胞菌的纖溶酶報道多見于海洋來源的假單胞菌,海洋微生物具有耐鹽、耐高壓、嗜低溫等其獨特的生理特性,是獲取新生物活性物質的又一重要來源。由青島海洋大學劉晨光等報道[26],從一株海洋假單胞菌中分離制備出一種具有纖溶活性的海洋假單胞菌堿性蛋白酶(MPAP),研究結果表明,該酶具有較強的直接溶解纖維蛋白的作用,靜脈用藥后,血栓形成時間明顯延長,血小板聚集率降低[27]。由此說明,MPAP具有明顯的抗血栓形成作用。但是對于該酶的結構、催化特征以及毒理藥效作用等還需進一步研究,為進一步開發該酶成為新的溶栓藥劑提供科學依據。
3.6 真菌類微生物產的纖溶酶
真菌類產纖溶酶的主要包括根霉、木霉、脈孢霉和幾種食用菌類。中華根霉12號是國內研究較早的產纖溶酶菌株,來源于傳統制酒行業的酒曲中,經過發酵可以產生類似于納豆激酶的絲氨酸蛋白酶;天津科技大學的孫哲等也從酒曲中篩選得到一株產纖溶酶的根霉,見鑒定為菊芋小孢根霉[28];米曲霉也是食品發酵重點重要菌株之一,高占爭等優化了米曲霉的產酶培養基,纖溶酶活性達到98.13 U/ml[29];天津科技大學的江潔等采用t-PA cDNA整合到里氏木霉的一株工程菌,發酵產t-PA進行了研究[30];脈孢霉是一種可以分解纖維素的真菌,在食用菌行業屬于污染菌株,齊齊哈爾大學的許愛清等對脈孢霉產纖溶酶菌株進行了誘變育種研究[31];沈陽藥科大學的吳黎誠篩選一株植物內生真菌鐮孢霉,產纖溶酶約28 KD,與其它纖溶酶無同源性,為絲氨酸蛋白酶和金屬蛋白酶[32];臺灣的張靜如研究了白僵菌所產纖溶酶[33];李華平等從冬蟲夏草中提取了纖溶酶CSP,分子量約31 KD,可以直接裂解纖維蛋白,不是纖溶酶原激活劑[34]。蛹蟲草產纖溶酶的研究最早見于齊齊哈爾大學的液體深層培養和江蘇農業科學院的蛹蟲草形態和分子鑒定[35-36],關于食用真菌類的有榆干離褶傘菌和黃綠蜜環菌產纖溶酶的報道[37-38]。
3.7 其它類別單株菌產纖溶酶
Vijayaraghavan P 從一株海洋希瓦氏菌Shewanellasp.IND20中分離出一株具有纖溶活性的酶[39]。沈陽藥科大學的李占強等報道了來源于海洋短小芽孢桿菌的纖溶酶[40]。蘭州生物制品研究所的趙紅等用產芽孢梭菌培養上清,純化得到了纖溶酶[41];肖璐等報道了一株產纖溶酶的藤黃微球菌,并且進行了基因克隆研究[42];西北農林科技大學的王高學等再青藏高原凍土內篩選得到一株溶栓菌,鑒定為金黃色桿菌[43],該株菌屬于放線菌門。重慶師范大學的和七一等篩選到一株產纖溶酶的糞腸球菌EF608,并進行了發酵工藝優化,纖溶酶活性達到1 833 U/ml[44]。
4 關于產纖溶酶的微生物分類比較
細菌和真菌中都有能產生纖溶酶的種類,特別是細菌中種類最多。由于細菌和真菌的多樣性,導致同一發酵物的纖溶酶也具有多樣性,具體詳細比較分別見表2和表3。

表2 產纖溶酶的細菌Table 2 Bacteria that produce fibrinolytic enzymes

表3 產纖溶酶的真菌Table 3 Fungi that produce fibrinolytic enzymes
5 展望
不論是來源于動物還是微生物的纖溶酶,都屬于絲氨酸蛋白酶,是為了自身獲得營養而進化的功能蛋白。例如蚯蚓是為了分解營養原料,水蛭、蝙蝠等是為了吸取不凝固的血液,微生物也是為了分解目的蛋白以獲得營養和自身繁殖,一些致病細菌或者蟲生真菌同樣是分解體細胞或蟲體蛋白的需要,所以一般也可以分泌纖溶酶。總觀各種來源的纖溶酶產物,雖然分子量大小稍有差異,從15~40 KD之間,并且大多數是絲氨酸蛋白酶類,針對基因序列比對(見圖2)和氨基酸序列比較(見圖3)后發現,不同纖溶酶基因序列雖然有很大差異,由于密碼子的簡并性,翻譯出來的氨基酸序列一致性比較高,說明絲氨酸蛋白酶活性功能區域是相對保守的,也符合生物繁衍過程中的趨同進化和趨異進化的理論。

圖3 不同菌株纖溶酶氨基酸序列比對Fig.3 Comparison of plasmin amino acid sequences of different strains
進一步推測:新發現微生物的菌株產纖溶酶的概率比較高,但是產新型纖溶酶的可能性很低。即便如此,由于微生物發酵具有周期短、成本低的特點,從不同環境中篩選微生物,并利用分子生物學和基因工程技術進行改造,生產不同作用方式的纖溶酶,一直還是生物和制藥行業關注的熱點。
