巨自噬與分子伴侶介導(dǎo)的自噬在饑餓誘導(dǎo)的Raji細胞中相繼順序被激活
2019-07-04 02:51:54李彩麗魏虎來
中國藥理學(xué)通報 2019年7期
關(guān)鍵詞:水平
李彩麗,辛 英,魏虎來,陳 靜,王 蓓,李 敏
(1.西北民族大學(xué)醫(yī)學(xué)院,甘肅 蘭州 730030;2.蘭州大學(xué)基礎(chǔ)醫(yī)學(xué)院,甘肅省新藥臨床前研究重點實驗室,甘肅 蘭州 730030)
自噬是進化上高度保守的溶酶體降解途徑,當(dāng)細胞在營養(yǎng)缺乏、缺氧、藥物等環(huán)境因素的影響下,細胞溶酶體對其內(nèi)部的細胞器、大分子物質(zhì)等內(nèi)容物進行“自我消化”的過程。細胞自噬所產(chǎn)生的物質(zhì)可以被細胞重新利用,以維持其生命活動[1]。自噬根據(jù)其發(fā)生的過程和降解底物的不同,分為巨自噬、分子伴侶介導(dǎo)的自噬和微自噬3種類型。巨自噬廣泛存在于真核細胞中,它的產(chǎn)生需要多種自噬相關(guān)基因參與,形成雙層膜結(jié)構(gòu)的吞噬泡,吞噬泡包裹細胞內(nèi)的可溶性蛋白和細胞器形成自噬體,繼而與溶酶體融合,將其內(nèi)容物進行降解加工[2-3]。分子伴侶介導(dǎo)的自噬(chaperone-mediated autophagy,CMA)是細胞自噬的一種特殊形式,也是哺乳動物所特有的自噬途徑。CMA的發(fā)生需要胞質(zhì)內(nèi)熱休克蛋白Hsc70與底物分子的特異氨基酸序列(如KFERQ-樣模體)識別結(jié)合,結(jié)合形成的復(fù)合物能被溶酶體相關(guān)膜蛋白(LAMP-2a)所識別,在溶酶體腔內(nèi)Hsc70的介導(dǎo)下,使底物去折疊并轉(zhuǎn)位至溶酶體內(nèi)進行降解[4-5]。Cuervo等[6-7]的研究表明,動物禁食3 d后,細胞內(nèi)CMA活性會明顯提高,胞質(zhì)內(nèi)含有KFERQ信號肽序列的蛋白質(zhì)明顯減少。這說明細胞在饑餓的不同階段所選擇的自噬途徑不同,它們順序被激活后,在功能上相互協(xié)調(diào)補充,共同參與細胞內(nèi)容物的降解,從而維持細胞的生命活動。然而,這種現(xiàn)象在腫瘤細胞中是否依然存在,目前尚無相關(guān)報道。本研究以Burkitt淋巴瘤Raji細胞為研究對象,旨在觀察體外培養(yǎng)的腫瘤細胞在饑餓誘導(dǎo)的不同階段,細胞巨自噬與CMA活性的變化情況,闡明巨自噬與CMA之間的關(guān)系,從而探索它們各自在腫瘤細胞應(yīng)對壓力刺激時的作用。
1 材料與方法
1.1 細胞株Burkitt淋巴瘤Raji細胞,購自中國科學(xué)院上海細胞生物學(xué)研究所。
1.2 試劑D-Hanks液(索萊寶公司);蛋白提取及定量試劑盒(碧云天公司);單丹黃酰尸胺(monodansylcadaverine,MDC)(Sigma公司);Hsc70一抗,Apitomics公司;LC3-Ⅰ/Ⅱ、Beclin-1、β-actin一抗,均購自CST公司; 溶酶體相關(guān)膜蛋白-2a(lysosome-associated membrane protein-2a,Lamp-2a)一抗,購自Abcam公司;紅外熒光標(biāo)記的二抗,購自美國LI-COR公司。
1.3 儀器CO2細胞培養(yǎng)箱(日本三洋公司);熒光顯微鏡(日本OLYMPUS公司);紅外熒光掃描成像系統(tǒng)(美國LI-COR公司);激光共聚焦顯微鏡(德國Leica公司)。
1.4 方法
1.4.1細胞培養(yǎng)與處理 采用D-Hanks緩沖液代替細胞培養(yǎng)液,饑餓誘導(dǎo)對數(shù)生長期的Raji細胞,分別收集饑餓誘導(dǎo)0、3、6、9、12 h的細胞進行檢測。
1.4.2MDC熒光染色觀察自噬體 在饑餓誘導(dǎo)不同時間的Raji細胞中,加入MDC貯存液,使其終濃度達到0.05 mmol·L-1,37 ℃避光孵育30 min,PBS洗滌2次,滴片,用激光共聚焦顯微鏡和熒光顯微鏡分別觀察細胞內(nèi)自噬體的形成。
1.4.3Western blot法檢測Beclin-1、LC3-Ⅰ/Ⅱ、Hsc70和Lamp-2a蛋白的表達 分別收集對照組以及饑餓誘導(dǎo)不同時間的Raji細胞,PBS洗滌,RIPA裂解液抽提細胞總蛋白,BCA試劑盒定量測定蛋白質(zhì)濃度,加入5×上樣緩沖液并混勻,每個樣品取50 μg蛋白質(zhì)上樣,SDS-PAGE電泳分離,轉(zhuǎn)膜、封閉,加入一抗室溫孵育1~2 h,洗滌后,加入二抗室溫孵育2 h,洗滌,用Odyssey紅外熒光掃描儀掃描并分析結(jié)果,每個實驗重復(fù)3次。
1.4.4統(tǒng)計學(xué)分析 實驗數(shù)據(jù)采用SPSS 13.0軟件進行分析處理,組間比較采用t檢驗。
2 結(jié)果
2.1 饑餓誘導(dǎo)不同時間Raji細胞巨自噬活性的改變采用D-Hanks緩沖液代替培養(yǎng)液饑餓誘導(dǎo)Raji細胞0~12 h,MDC染色后,分別用激光共聚焦顯微鏡和熒光顯微鏡觀察Raji細胞內(nèi)自噬體的形成情況。如Fig 1所示,在饑餓誘導(dǎo)的最初幾小時,MDC熒光強度逐漸增加,在6 h達到峰值,隨后逐漸下降到基礎(chǔ)水平。
2.2 饑餓誘導(dǎo)不同時間巨自噬相關(guān)蛋白的表達水平Western blot檢測饑餓誘導(dǎo)不同階段,Raji細胞內(nèi)巨自噬標(biāo)志性蛋白LC3-Ⅰ/Ⅱ和Beclin-1的表達水平。如Fig 2所示,LC3-Ⅱ和Beclin-1的表達水平在饑餓誘導(dǎo)的最初幾小時均逐漸升高,6 h時達到峰值,隨后又逐漸下降。
2.3 饑餓誘導(dǎo)不同時間細胞內(nèi)CMA相關(guān)蛋白的表達水平Hsc-70和Lamp-2a是參與CMA的兩種重要物質(zhì),分子伴侶Hsc70能特異性識別帶有KFERQ序列的底物蛋白,促進底物去折疊并與LAMP-2a結(jié)合,繼而進入溶酶體腔被降解[8-9]。因此,Hsc70和LAMP-2a是檢測CMA活性的重要標(biāo)志物。Western blot檢測饑餓誘導(dǎo)0~12 h細胞內(nèi)CMA的水平,發(fā)現(xiàn)Hsc-70和Lamp-2a的表達并未像巨自噬那樣在饑餓誘導(dǎo)的早期(3~6 h)明顯升高,而是在饑餓誘導(dǎo)6 h之后才明顯增高,12 h時達到峰值(Fig 3)。
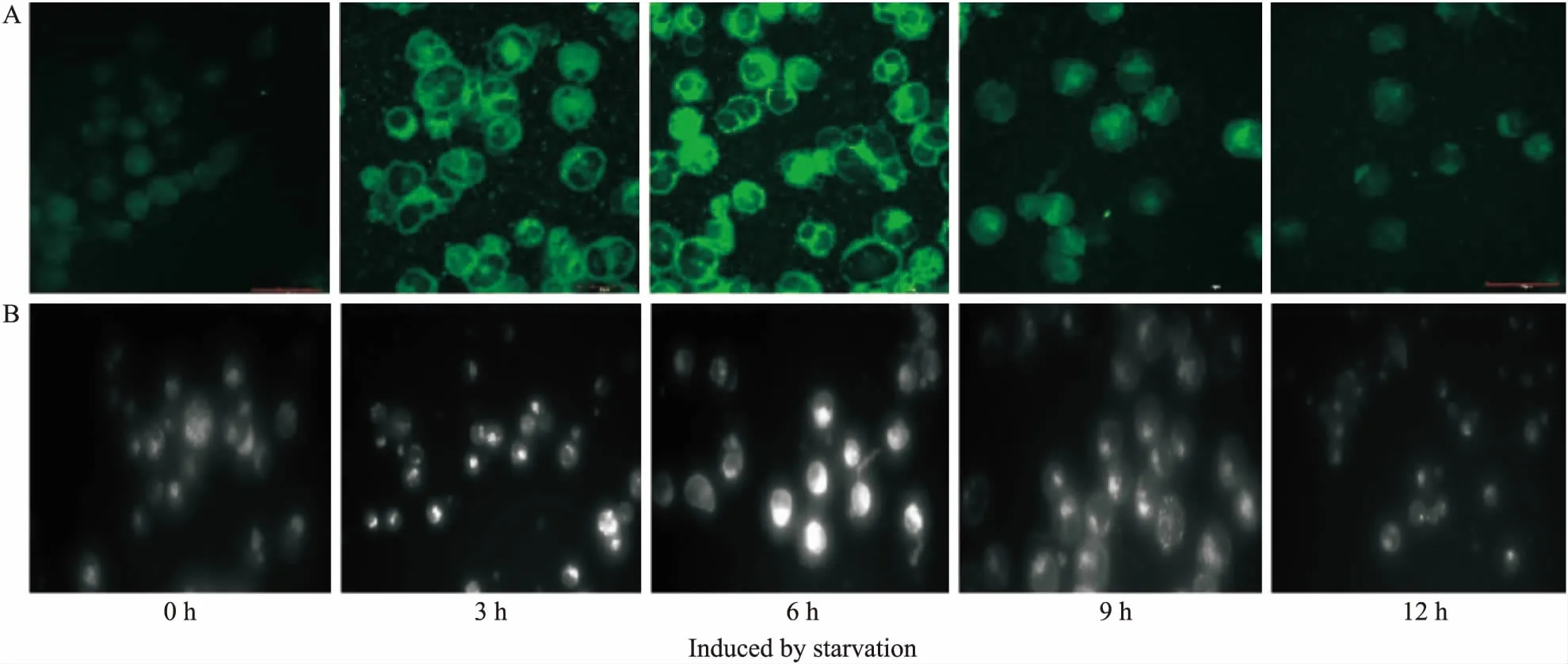
Fig 1 The fluorescence intensity of MDC in Raji cells induced by starvation(×100)
A:Detection results of confocal laser-scanning microscopy;B: The fluorescence intensity of MDC was analysed by fluorescence microscopy.
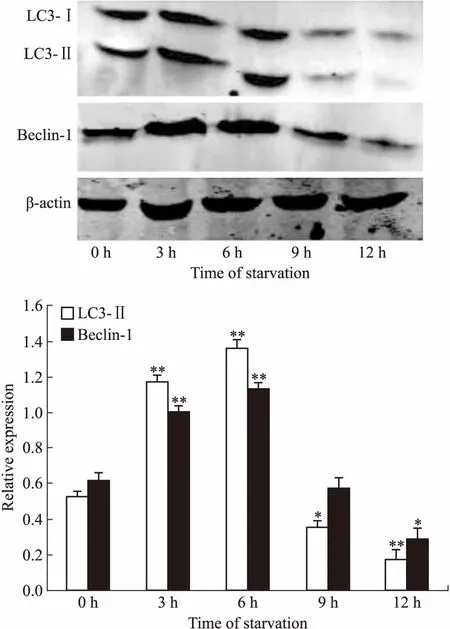
Fig 2 Macroautophagy-related protein expression in starvation-induced Raji cells
*P<0.05,**P<0.01vscontrol group(0 h)
3 討論
CMA與巨自噬作為細胞自噬的兩種不同類型,其在腫瘤治療中的作用越來越受到人們的關(guān)注。盡管它們在發(fā)生的先后順序、對底物的選擇性以及調(diào)控機制等很多方面不同,然而它們在維持機體內(nèi)環(huán)境穩(wěn)定方面卻有著緊密的聯(lián)系。有研究發(fā)現(xiàn),巨自噬與CMA在正常細胞中存在相互協(xié)調(diào)補充的關(guān)系,即當(dāng)巨自噬活性下降或被抑制時,CMA就會被活化,從而補償巨自噬的部分功能[10-13],這說明它們并非獨立存在的兩條途徑,而是存在著必然的聯(lián)系。然而,這種現(xiàn)象是否在腫瘤細胞中依然存在,目前尚無相關(guān)報道。
本研究通過饑餓誘導(dǎo)Burkitt淋巴瘤Raji細胞,觀察細胞內(nèi)巨自噬和CMA活性的變化,探究腫瘤細胞受到壓力刺激時,巨自噬與CMA之間的關(guān)系。本研究發(fā)現(xiàn),Raji細胞在D-Hanks緩沖液中分別培養(yǎng)0、3、6、9、12 h,細胞巨自噬活性在饑餓初期(3~6 h)迅速增加并達到峰值,與對照組(0 h)相比較,胞質(zhì)內(nèi)MDC染色陽性的自噬體數(shù)量,以及LC3-Ⅱ和Beclin-1表達水平在饑餓6 h時均達到高峰,與此相反,CMA相關(guān)分子Hsc70和LAMP-2a的表達水平在此時并未見明顯上調(diào)。然而,隨著饑餓時間的延長,細胞巨自噬活性逐漸降低,并伴隨出現(xiàn)CMA活性的逐漸增加,當(dāng)饑餓12 h時,巨自噬活性基本達到對照組細胞水平,而CMA活性此時卻達到了峰值。由此證明,巨自噬是細胞應(yīng)對饑餓這種壓力刺激時的一種快速而短暫的反應(yīng),細胞首先通過巨自噬降解胞質(zhì)內(nèi)生物大分子,實現(xiàn)物質(zhì)的循環(huán)再利用,為處于饑餓狀態(tài)的細胞提供首批氨基酸和能量等,以維持生存的需要。隨之巨自噬水平的降低可有效避免一些重要蛋白質(zhì)的非選擇性和無序性降解,與此同時,CMA的活化通過選擇性降解一些非必需的蛋白質(zhì),進一步為細胞生存提供能源物質(zhì)。
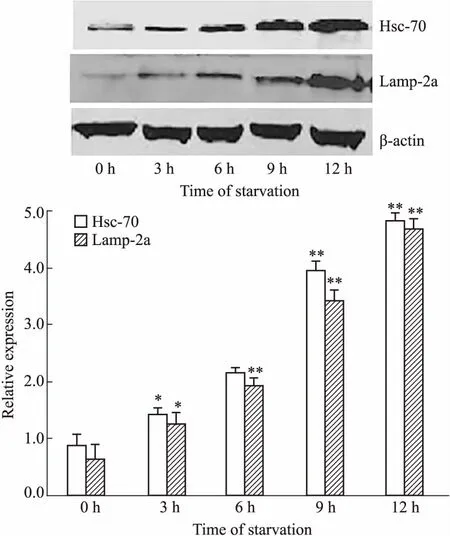
Fig 3 CMA-related protein expression in starvation-induced Raji cells
*P<0.05,**P<0.01vscontrol group(0 h)
本研究證實,Burkitt淋巴瘤Raji細胞在接受饑餓刺激時,巨自噬與CMA之間存在相互協(xié)調(diào)和補充的關(guān)系,它們并非同步而是相繼被激活,巨自噬的發(fā)生先于CMA,高巨自噬水平會抑制CMA的活化,只有當(dāng)巨自噬水平下降時,才會出現(xiàn)CMA活性的增加。這一發(fā)現(xiàn)將有助于更好地理解腫瘤細胞中不同自噬途徑的特點、相互之間的關(guān)系,以及在應(yīng)對壓力刺激時的應(yīng)答機制。
猜你喜歡
美與時代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30