肥胖患者脂肪組織STAU1表達(dá)變化及其在脂肪間充質(zhì)干細(xì)胞成脂分化中的作用
2019-06-18 02:35:22孟軒羽梁小弟努爾比耶努爾麥麥提焦誼劉杰胡婷婷高佳樂(lè)徐尤宗盛張玲關(guān)亞群
山東醫(yī)藥 2019年14期
關(guān)鍵詞:水平
孟軒羽,梁小弟,努爾比耶·努爾麥麥提,焦誼,劉杰,胡婷婷,高佳樂(lè),徐尤宗盛,張玲,關(guān)亞群
(新疆醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院,烏魯木齊830011)
肥胖是引起2型糖尿病、高血壓、非酒精性脂肪肝等代謝性疾病的重要原因之一[1~4],脂肪細(xì)胞的異常分化和過(guò)度增殖是導(dǎo)致肥胖的細(xì)胞生物學(xué)基礎(chǔ)。脂肪細(xì)胞分化涉及一個(gè)復(fù)雜的調(diào)控網(wǎng)絡(luò),包括表觀遺傳學(xué)、轉(zhuǎn)錄水平、轉(zhuǎn)錄后水平、翻譯及翻譯后水平[5];在轉(zhuǎn)錄水平中,過(guò)氧化物酶體增殖劑激活受體(PPARγ)不僅是調(diào)控脂肪組織中前體脂肪細(xì)胞分化的關(guān)鍵因子,而且在調(diào)節(jié)脂類(lèi)代謝中扮演重要的角色[6]。在轉(zhuǎn)錄后水平中,RNA結(jié)合蛋白起著重要的調(diào)節(jié)作用[7]。Cho等[8]發(fā)現(xiàn),雙鏈RNA結(jié)合蛋白STAU1在小鼠脂肪細(xì)胞3T3-L1的分化過(guò)程中高表達(dá)。前期我們通過(guò)生物信息學(xué)方法預(yù)測(cè)發(fā)現(xiàn),STAU1可以與PPARγ結(jié)合。2018年7~9月,我們?cè)谌酥窘M織及人脂肪間充質(zhì)干細(xì)胞(hADSCs)成脂分化中檢測(cè)STAU1的表達(dá)水平,并于hADSCs中敲低STAU1表達(dá)后檢測(cè)細(xì)胞成脂情況及PPARγ表達(dá)水平變化,初步探討STAU1在人脂肪組織及hADSCs成脂分化中的可能作用。
1 材料與方法
1.1 主要試劑與儀器 低糖培養(yǎng)基(DMEM)、胎牛血清(FBS)購(gòu)自Hyclone;無(wú)血清培養(yǎng)基(Opti-MEM)購(gòu)自Gibco-BRL;LipofectamineTM3000轉(zhuǎn)染試劑、TRIzol RNA提取試劑購(gòu)自Invitrogen;SYBR Green Real time PCR試劑盒購(gòu)自Takala;RevertAid First Strand cDNA Synthesis Kit逆轉(zhuǎn)錄試劑盒購(gòu)自Thermo Scientific;Anti-actin、Anti-STAU1、Anti-PPARγ抗體購(gòu)自Abcam;STAU1 siRNA購(gòu)自廣州瑞博。熒光定量PCR儀(ABI 7500)核酸微量檢測(cè)儀購(gòu)自NanoDrop;低溫高速離心機(jī)購(gòu)自Thermo,熒光倒置顯微鏡購(gòu)自Leica;全波長(zhǎng)酶標(biāo)儀、凝膠成像儀購(gòu)自Bio-Rad;電熱恒溫培養(yǎng)箱購(gòu)自Thermo。
1.2 實(shí)驗(yàn)方法
1.2.1 人脂肪組織的獲取與臨床資料收集 選取2016年9月~2018年5月新疆維吾爾自治區(qū)人民醫(yī)院慢性膽囊結(jié)石和腹股溝疝擇期手術(shù)患者56例,均無(wú)動(dòng)脈或外周血管疾病、甲狀腺功能紊亂、急性炎癥性疾病、創(chuàng)傷和其他需要緊急治療的病癥、惡性腫瘤、酒精或藥物濫用史者,術(shù)中留取網(wǎng)膜和皮下脂肪組織。將患者根據(jù)BMI分為肥胖組(BMI≥28 kg/m2)27例和非肥胖組(BMI<24 kg/m2)29例,兩組年齡、BMI、腰圍、臀圍、血壓、血糖、血脂比較,見(jiàn)表1。本研究均經(jīng)醫(yī)院倫理委員會(huì)批準(zhǔn),患者均簽署知情同意書(shū)。
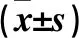
表1 兩組年齡、BMI、腰圍、臀圍、血壓、血糖、血脂比較
注:與非肥胖組比較,*P<0.05。
1.2.2 hADSCs的獲取 取人網(wǎng)膜脂肪組織約2 g,用含1 000 U/mL青霉素和鏈霉素的PBS清洗3遍;用眼科剪將脂肪組織剪至1 mm3大小,在37 ℃水浴中與1 mg/mL的Ⅰ型膠原酶一起溫育60 min,輕微搖晃直至看不到組織塊為止。加入20 mL PBS終止消化,將組織液以600 r/min離心10 min。棄去上清液后,將沉淀重懸于含有10% FBS和1 000 U/mL青霉素和鏈霉素的低糖DMEM中。將細(xì)胞懸浮液接種在10 cm細(xì)胞培養(yǎng)皿上,在37 ℃培養(yǎng)箱中用5% CO2培養(yǎng);48 h換1次液,直至細(xì)胞融合率為80%,用胰酶消化傳代。傳代后收集細(xì)胞懸液,分為陰性對(duì)照及同型對(duì)照,分別加單克隆抗體CD44-FITC、CD105-PE在4 ℃冰箱避光孵育30 min;用PBS清洗5遍,洗掉多余的抗體后上流式細(xì)胞儀檢測(cè)細(xì)胞表面抗原。結(jié)果顯示,間充質(zhì)干細(xì)胞標(biāo)記抗原CD44、CD105呈陽(yáng)性表達(dá),表明分離的細(xì)胞為hADSCs。
1.2.3 hADSCs成脂分化誘導(dǎo)與甘油三酯形成情況觀察 當(dāng)細(xì)胞融合率達(dá)85%時(shí),將含10%FBS、1 μmol/L地塞米松、10 μmol/L胰島素、0.5 mmol/L 3-異丁基-1-甲基黃嘌呤和200 μmol/L吲哚美辛的成脂誘導(dǎo)液處理hADSCs;每3 d更換培養(yǎng)基1次,持續(xù)8 d。分別于0、2、4、6、8 d,取上述細(xì)胞樣品;PBS清洗后以4%多聚甲醛固定30 min,用油紅O染料染色25 min。PBS清洗3遍后,加入ddH2O放在倒置顯微鏡下觀察細(xì)胞成脂情況;最后用異丙醇洗脫培養(yǎng)皿提取油紅O染料,在波長(zhǎng)520 nm下測(cè)量光密度,以此表示甘油三酯生成量。
1.2.4 脂肪組織STAU1基因和hADSCs成脂分化中STAU1、PPARγ基因表達(dá)檢測(cè) 采用RT-qPCR法。PBS洗滌3.5 cm培養(yǎng)皿3次后(脂肪組織用研缽碾成粉末),加入TRIzol 1 mL吹打勻漿細(xì)胞和組織;采用異丙醇氯仿方法提取細(xì)胞及組織內(nèi)RNA,通過(guò)分光光度法測(cè)量RNA濃度及A260/A280,確保RNA的質(zhì)量。使用Revert Aid First Strand cDNA Synthesis試劑盒將RNA逆轉(zhuǎn)錄為cDNA后,在ABI 7500系統(tǒng)下使用SYBR Select Master Mix試劑進(jìn)行PCR擴(kuò)增。STAU1基因上游引物5′-TGCTGGGGTTTGTTGTGATG-3′、下游引物5′-TGAGTGAGCCAGCTTGAGAA-3′,PPARγ基因上游引物5′-AAATGCGAGAGTGGGAAGGA-3′、下游引物5′-AGGGTCTTGCTATGTTGCCT-3′,由上海生工生物有限公司設(shè)計(jì)并合成。以HSP90為內(nèi)參,用2-ΔΔCt法計(jì)算STAU1基因相對(duì)表達(dá)量。
1.2.5 脂肪組織STAU1蛋白和hADSCs成脂分化中STAU1、PPARγ蛋白表達(dá)檢測(cè) 采用Western blotting法。將hADSCs細(xì)胞用PBS清洗3遍后(網(wǎng)膜脂肪組織研磨成粉末),在RIPA中裂解后用BCA工作液對(duì)蛋白進(jìn)行定量;在10%的SDS-PAGE膠上樣電泳后轉(zhuǎn)移至PVDF膜,在5% BSA中室溫孵育2 h;將膜和一抗Anti-STAU1、Anti-HSP90、Anti-PPARγ(1∶1 000)置于5 mL一抗稀釋緩沖液中,4 ℃搖床上孵育過(guò)夜。用15 mL TBST洗滌3次,每次15 min;將二抗(1∶5 000)HRP-Anti-rabbit室溫孵育1 h,再用15 mL TBST洗滌3次,每次15 min。ECL底物進(jìn)行顯色,以Image Lab軟件進(jìn)行光密度分析。以HSP90為內(nèi)參,用目的蛋白與內(nèi)參吸光度比值表示目的蛋白相對(duì)表達(dá)量。
1.2.6 敲低STAU1對(duì)hADSCs中甘油三酯生成及PPARγ表達(dá)影響的觀察 將4×105個(gè)細(xì)胞接種于3.5 cm皿中,待融合率為60%時(shí),加入基礎(chǔ)培養(yǎng)基2 mL饑餓8 h后;將細(xì)胞分為3組,干擾1、2組分別轉(zhuǎn)染siRNA STAU1-homo-718、STAU1-homo-923,對(duì)照組不轉(zhuǎn)染。加無(wú)血清培養(yǎng)基Opti-MEM 250 μL稀釋10 μL LipofectamineTM3000,室溫孵育5 min;加入5 μL siRNA用250 μL Opti-MEM進(jìn)行稀釋?zhuān)瑢⒅|(zhì)體LipofectamineTM3000與siRNA混合液室溫孵育30 min;加入培養(yǎng)皿中在37 ℃、5% CO2的培養(yǎng)箱中孵育1 h后換成DMEM低糖完全培養(yǎng)基,每2 d換液1次。轉(zhuǎn)染后2、4 d,分別檢測(cè)甘油三酯生成情況、STAU1和PPARγ基因及蛋白質(zhì)表達(dá)水平,方法分別同1.2.3、1.2.4、1.2.5。siRNA由廣州博瑞設(shè)計(jì)并合成,引物序列見(jiàn)表2。

表2 STAU1 siRNA引物序列

2 結(jié)果
2.1 肥胖組與非肥胖組脂肪組織中STAU1基因及蛋白表達(dá)情況 肥胖組網(wǎng)膜脂肪組織中STAU1基因相對(duì)表達(dá)量高于非肥胖組(P<0.05),兩組皮下脂肪組織中STAU1基因相對(duì)表達(dá)量差異無(wú)統(tǒng)計(jì)學(xué)意義,見(jiàn)表3。肥胖組網(wǎng)膜脂肪組織中STAU1蛋白相對(duì)表達(dá)量為2.68±0.28,高于非肥胖組的1.32±0.36(P<0.05)。
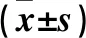
表3 肥胖組與非肥胖組脂肪組織中STAU1基因表達(dá)比較
注:與非肥胖組同型脂肪組織比較,*P<0.05。
2.2 肥胖組網(wǎng)膜脂肪組織中STAU1基因表達(dá)水平與患者臨床特征的關(guān)系 相關(guān)性分析顯示,肥胖組網(wǎng)膜脂肪組織中STAU1基因表達(dá)水平與患者的腰圍、臀圍、低密度脂蛋白、高密度脂蛋白、空腹血糖無(wú)明顯相關(guān)性(r分別為0.169、0.176、0.173、0.164、0.171,P均>0.05),與BMI、甘油三酯呈正相關(guān)(r分別為0.777、0.795,P均<0.01)。
2.3 hADSCs成脂分化過(guò)程中甘油三酯生成及STAU1、PPARγ表達(dá)水平變化 隨著誘導(dǎo)分化天數(shù)增加,hADSCs中甘油三酯生成增多,STAU1、PPARγ表達(dá)水平逐漸升高(P均<0.05)。見(jiàn)表4。
表4 hADSCs成脂分化過(guò)程中甘油三酯生成及STAU1、PPARγ表達(dá)水平變化
注:與成脂分化誘導(dǎo)0 d時(shí)比較,*P<0.05。
2.4 敲低STAU1對(duì)hADSCs中甘油三酯生成及PPARγ表達(dá)的影響 與對(duì)照組比較,干擾1、2組轉(zhuǎn)染2、4 d時(shí)hADSCs中STAU1表達(dá)降低,甘油三酯生成及PPARγ表達(dá)減少(P均<0.05)。見(jiàn)表5。
表5 各組hADSCs中甘油三酯生成及STAU1、PPARγ表達(dá)比較
注:與對(duì)照組同時(shí)點(diǎn)比較,*P<0.05。
3 討論
隨著社會(huì)發(fā)展及生活方式的改變,肥胖得到人們?cè)絹?lái)越多的關(guān)注,其發(fā)病率在全球范圍內(nèi)迅速上升[9~12]。研究發(fā)現(xiàn),脂肪細(xì)胞的異常分化是導(dǎo)致肥胖的重要原因之一。因此,研究脂肪細(xì)胞的分化及其機(jī)制對(duì)預(yù)防和治療肥胖及相關(guān)疾病具有重要意義。脂肪細(xì)胞分化可以分為兩個(gè)階段:第一個(gè)階段為多潛能的干細(xì)胞轉(zhuǎn)變?yōu)榍绑w脂肪細(xì)胞;第二個(gè)階段為前體脂肪細(xì)胞終末分化為成熟的脂肪細(xì)胞[13]。脂肪細(xì)胞分化存在復(fù)雜的調(diào)控網(wǎng)絡(luò),如PPARγ和CEBPα轉(zhuǎn)錄因子介導(dǎo)的轉(zhuǎn)錄水平的調(diào)節(jié)[14]。PPARγ通過(guò)調(diào)控下游與脂代謝相關(guān)基因的表達(dá),促進(jìn)脂肪細(xì)胞的分化、增加脂肪細(xì)胞的數(shù)量,對(duì)脂肪的生成、轉(zhuǎn)運(yùn)及脂肪的貯存和氧化等都起著重要作用,是脂肪細(xì)胞形成和代謝的關(guān)鍵調(diào)節(jié)因子[15]。另外,轉(zhuǎn)錄后水平也在脂肪細(xì)胞分化過(guò)程中發(fā)揮著重要作用,其中RNA結(jié)合蛋白可以參與RNA前體的剪接、RNA的細(xì)胞定位、RNA穩(wěn)定性等多種轉(zhuǎn)錄后調(diào)控[16]。STAU1是重要的RNA結(jié)合蛋白,又稱STAU1人源全長(zhǎng)重組蛋白。Cho等[17]報(bào)道,在前體脂肪細(xì)胞3T3-L1分化中,RNA結(jié)合蛋白STAU1可以通過(guò)與Kruppel樣因子2(KLF2)基因上的局部雙連結(jié)合,影響KLF2基因穩(wěn)定性,使KLF2下游靶基因PPARγ表達(dá)水平升高,進(jìn)而促進(jìn)3T3-L1細(xì)胞內(nèi)甘油三脂的生成。
本實(shí)驗(yàn)在hADSCs成脂分化過(guò)程中,STAU1、PPARγ表達(dá)水平隨著誘導(dǎo)分化天數(shù)增加而增加,油紅O染色結(jié)果顯示成脂加快。生物信息學(xué)方法預(yù)測(cè)顯示STAU1可以與PPARγ結(jié)合,而PPARγ是脂肪細(xì)胞分化重要的轉(zhuǎn)錄因子。本研究結(jié)果顯示,在脂肪細(xì)胞中敲低STAU1后PPARγ的蛋白及基因水平降低,推測(cè)STAU1可能通過(guò)上調(diào)PPARγ的表達(dá)水平進(jìn)而加速甘油三酯的生成,此外,在人脂肪組織中發(fā)現(xiàn)STAU1基因和蛋白水平在肥胖患者網(wǎng)膜脂肪組織中表達(dá)升高,差異有統(tǒng)計(jì)學(xué)意義,而在皮下脂肪組織中無(wú)統(tǒng)計(jì)學(xué)意義。Serralde-Zúniga等[18]研究發(fā)現(xiàn),脂肪組織在脂代謝和病理生理方面有明顯的部位特異性,網(wǎng)膜脂肪的積聚較皮下脂肪對(duì)人體危害更加明顯。因此,STAU1在網(wǎng)膜脂肪組織高表達(dá)可能與內(nèi)臟肥胖導(dǎo)致的代謝性疾病相關(guān)。相關(guān)性分析發(fā)現(xiàn),網(wǎng)膜脂肪組織中STAU1基因表達(dá)與肥胖組BMI和甘油三酯呈正相關(guān)。甘油三酯是組成脂質(zhì)的主要成分,脂肪酸作為合成甘油三酯的直接底物,脂肪酸合成增多直接加速了甘油三酯的合成;PPARγ可以通過(guò)調(diào)節(jié)脂肪酸合成酶的速率,加速脂肪酸合成的進(jìn)程,進(jìn)而增加了甘油三酯的合成[19,20]。這提示STAU1在脂肪組織及脂肪細(xì)胞中可能通過(guò)上調(diào)PPARγ的表達(dá),促進(jìn)甘油三酯的生成,從而導(dǎo)致肥胖的發(fā)生。
綜上所述,雙鏈RNA結(jié)合蛋白STAU1,在人網(wǎng)膜脂肪組織及hADSCs成脂分化過(guò)程中高表達(dá),STAU1可通過(guò)調(diào)節(jié)PPARγ表達(dá)水平來(lái)影響人體脂肪的形成,從而在肥胖的發(fā)生發(fā)展中起重要作用,但具體調(diào)控機(jī)制還有待進(jìn)一步研究。
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問(wèn)題研究(2012年1期)2012-03-25 09:54:45
中國(guó)火炬(2010年12期)2010-07-25 13:26:22
中國(guó)火炬(2010年8期)2010-07-25 11:34:30

- 山東醫(yī)藥的其它文章
- PI3K抑制劑對(duì)2型糖尿病小鼠腦組織中乙酰膽堿酯酶表達(dá)的影響
- 不同方法溶解20%甘露醇注射液結(jié)晶的質(zhì)量評(píng)價(jià)
- 高血壓腦出血術(shù)后應(yīng)用高壓氧治療對(duì)患者血清MMP-9、ICAM-1、NSE、hs-CRP水平的影響
- 腹腔鏡手術(shù)聯(lián)合GnRH-a治療子宮內(nèi)膜異位癥臨床效果及對(duì)患者卵巢功能的影響
- 波形蛋白、鋅指蛋白在非小細(xì)胞肺癌組織中的表達(dá)變化及其相互關(guān)系
- 血液透析聯(lián)合腹膜透析治療對(duì)尿毒癥患者營(yíng)養(yǎng)狀況、心腎功能及并發(fā)癥的影響